

A temporal chromatin signature in human embryonic stem cells identifies regulators of cardiac development
- PMID: 22981225
- PMCID: PMC3462257
- DOI: 10.1016/j.cell.2012.08.027
A temporal chromatin signature in human embryonic stem cells identifies regulators of cardiac development
Abstract
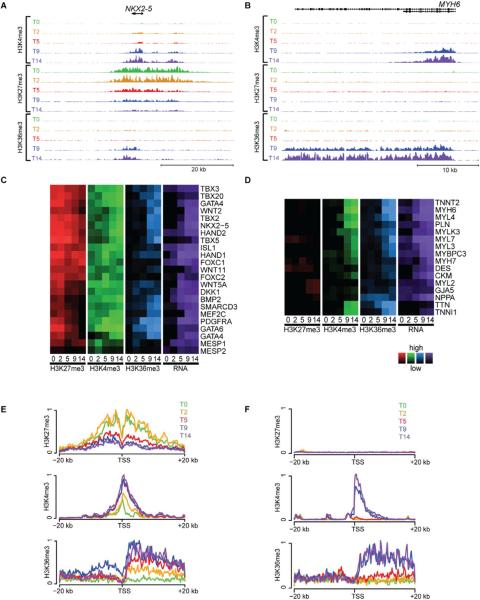
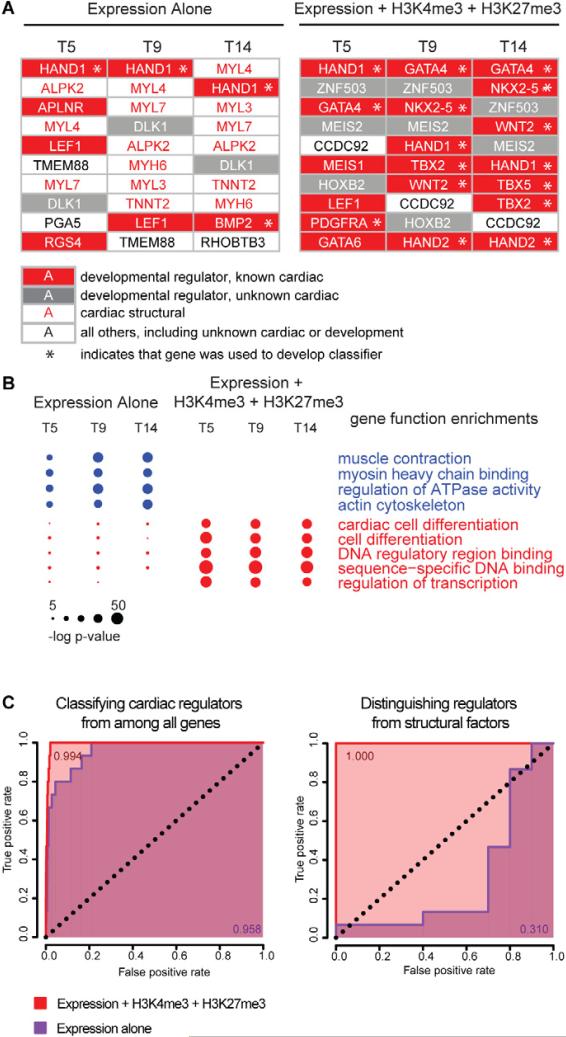
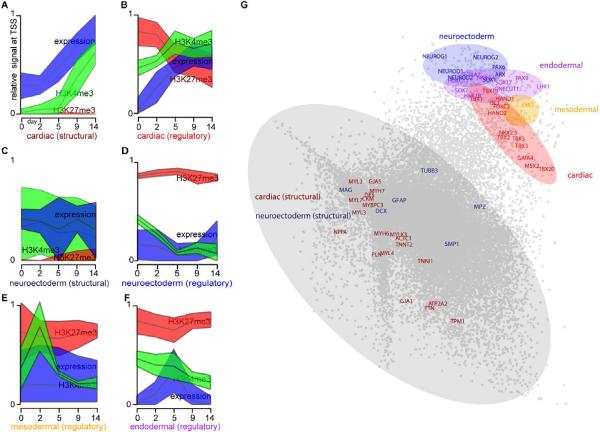
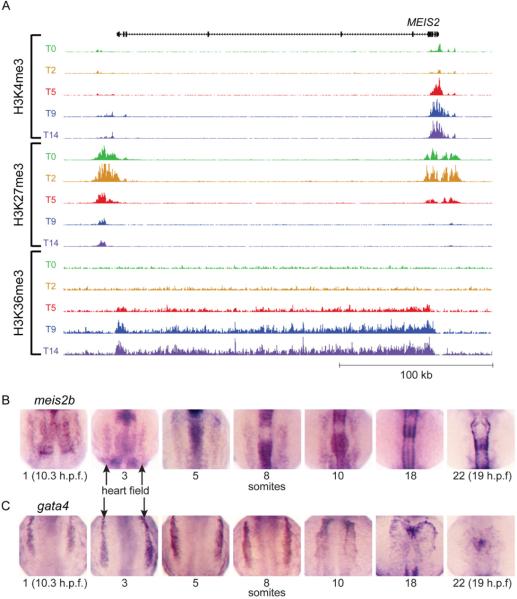
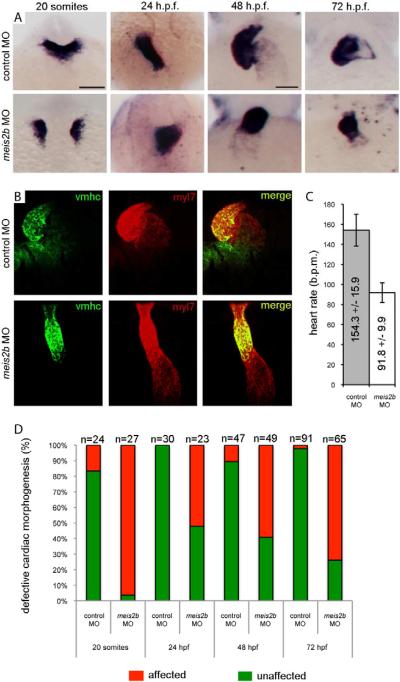
Directed differentiation of human embryonic stem cells (ESCs) into cardiovascular cells provides a model for studying molecular mechanisms of human cardiovascular development. Although it is known that chromatin modification patterns in ESCs differ markedly from those in lineage-committed progenitors and differentiated cells, the temporal dynamics of chromatin alterations during differentiation along a defined lineage have not been studied. We show that differentiation of human ESCs into cardiovascular cells is accompanied by programmed temporal alterations in chromatin structure that distinguish key regulators of cardiovascular development from other genes. We used this temporal chromatin signature to identify regulators of cardiac development, including the homeobox gene MEIS2. Using the zebrafish model, we demonstrate that MEIS2 is critical for proper heart tube formation and subsequent cardiac looping. Temporal chromatin signatures should be broadly applicable to other models of stem cell differentiation to identify regulators and provide key insights into major developmental decisions.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Maps for changing landscapes: viewing epigenomic signatures through differentiation.Cell Stem Cell. 2012 Nov 2;11(5):581-2. doi: 10.1016/j.stem.2012.10.009. Cell Stem Cell. 2012. PMID: 23122282
References
-
- Azuara V, Perry P, Sauer S, Spivakov M, Jørgensen HF, John RM, Gouti M, Casanova M, Warnes G, Merkenschlager M, et al. Chromatin signatures of pluripotent cell lines. Nat Cell Biol. 2006;8:532–538. - PubMed
-
- Bannister AJ, Schneider R, Myers FA, Thorne AW, Crane-Robinson C, Kouzarides T. Spatial distribution of di- and tri-methyl lysine 36 of histone H3 at active genes. J Biol Chem. 2005;280:17732–17736. - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
-
- Boyer LA, Plath K, Zeitlinger J, Brambrink T, Medeiros LA, Lee TI, Levine SS, Wernig M, Tajonar A, Ray MK, et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature. 2006;441:349–353. - PubMed
-
- Bu L, Jiang X, Martin-Puig S, Caron L, Zhu S, Shao Y, Roberts DJ, Huang PL, Domian IJ, Chien KR. Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature. 2009;460:113–117. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
- P01 HL094374/HL/NHLBI NIH HHS/United States
- U01 HL100405/HL/NHLBI NIH HHS/United States
- U54HG004592/HG/NHGRI NIH HHS/United States
- R01 HL084642/HL/NHLBI NIH HHS/United States
- R01 HL064387/HL/NHLBI NIH HHS/United States
- P01 GM081619/GM/NIGMS NIH HHS/United States
- F30 HL095343/HL/NHLBI NIH HHS/United States
- U54 HG004592/HG/NHGRI NIH HHS/United States
- R03 AR057477/AR/NIAMS NIH HHS/United States
- R01 HL64387/HL/NHLBI NIH HHS/United States
- R90HG004152/HG/NHGRI NIH HHS/United States
- P01 GM081719/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
