

Epigenomic annotation of enhancers predicts transcriptional regulators of human neural crest
- PMID: 22981823
- PMCID: PMC3751405
- DOI: 10.1016/j.stem.2012.07.006
Epigenomic annotation of enhancers predicts transcriptional regulators of human neural crest
Abstract
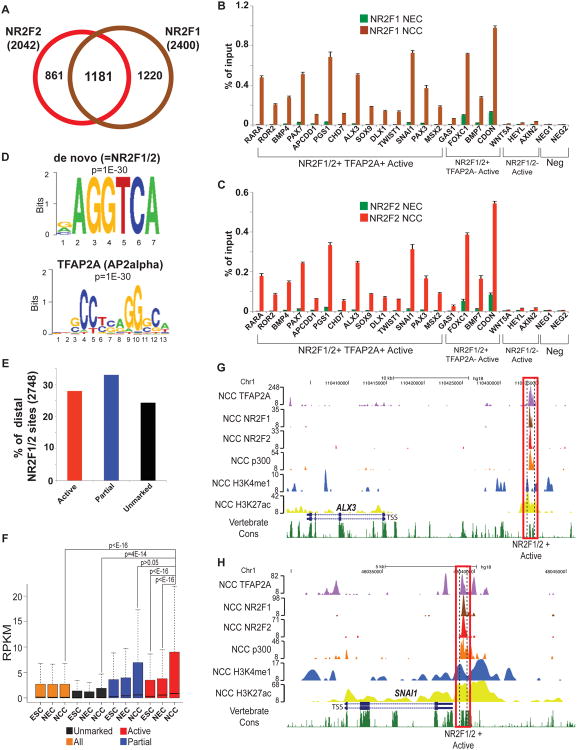
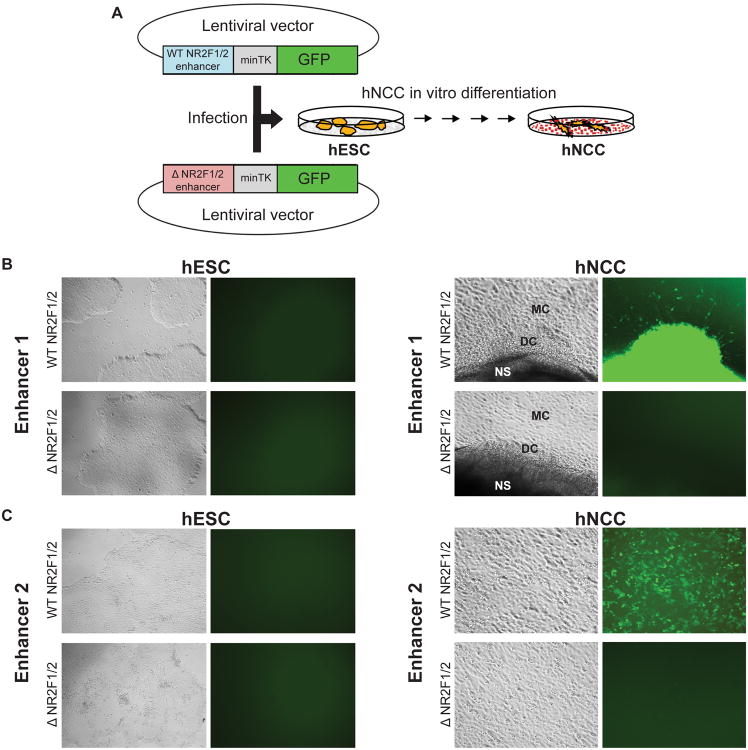
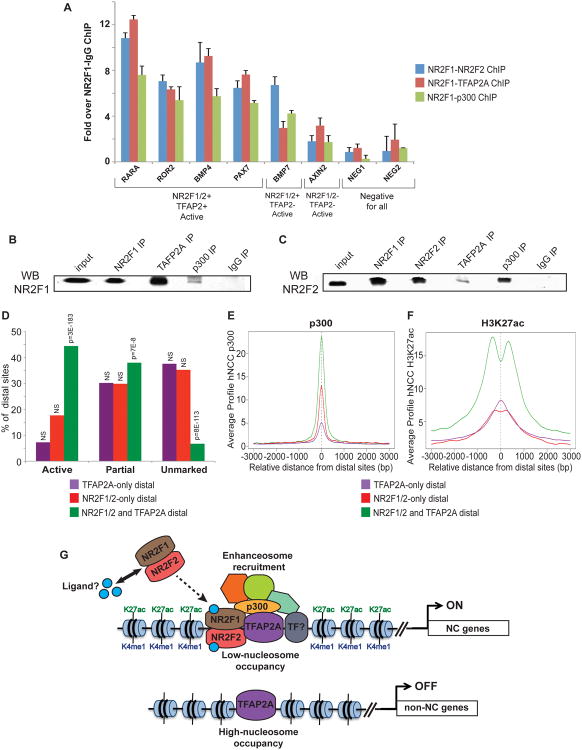
Neural crest cells (NCC) are a transient, embryonic cell population characterized by unusual migratory ability and developmental plasticity. To annotate and characterize cis-regulatory elements utilized by the human NCC, we coupled a hESC differentiation model with genome-wide profiling of histone modifications and of coactivator and transcription factor (TF) occupancy. Sequence analysis predicted major TFs binding at epigenomically annotated hNCC enhancers, including a master NC regulator, TFAP2A, and nuclear receptors NR2F1 and NR2F2. Although many TF binding events occur outside enhancers, sites coinciding with enhancer chromatin signatures show significantly higher sequence constraint, nucleosomal depletion, correlation with gene expression, and functional conservation in NCC isolated from chicken embryos. Simultaneous co-occupancy by TFAP2A and NR2F1/F2 is associated with permissive enhancer chromatin states, characterized by high levels of p300 and H3K27ac. Our results provide global insights into human NC chromatin landscapes and a rich resource for studies of craniofacial development and disease.
Copyright © 2012 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
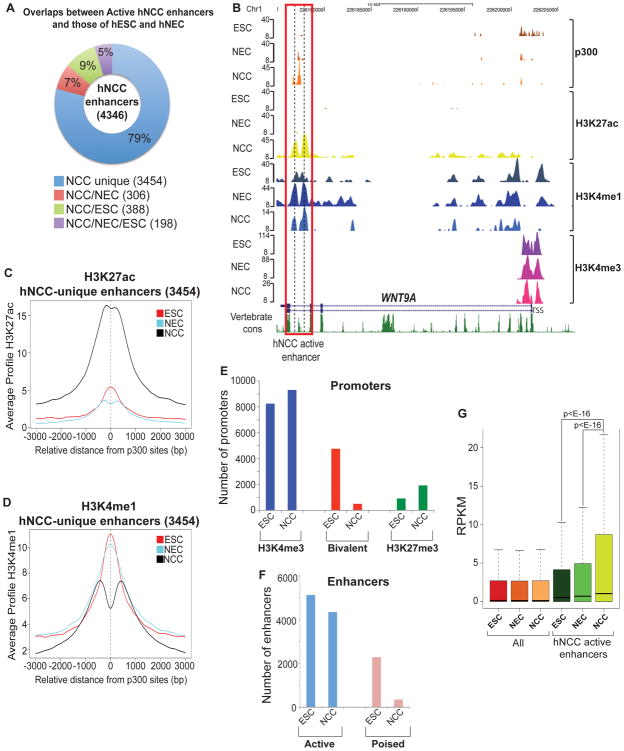
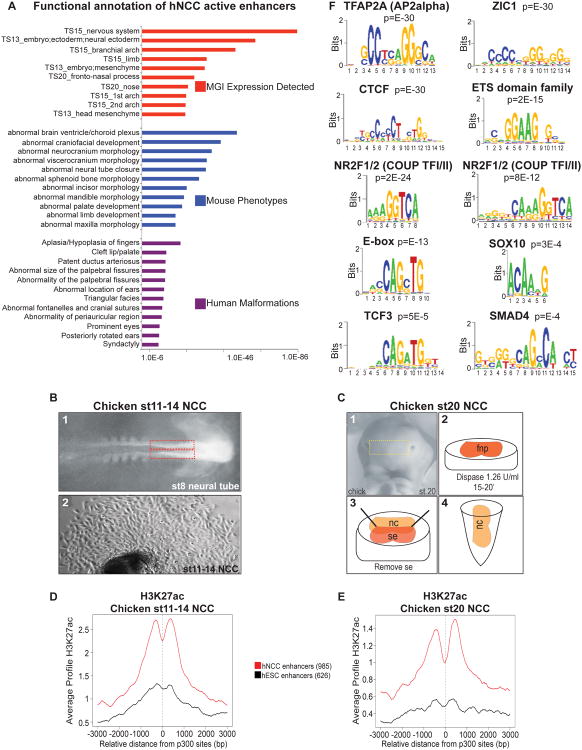
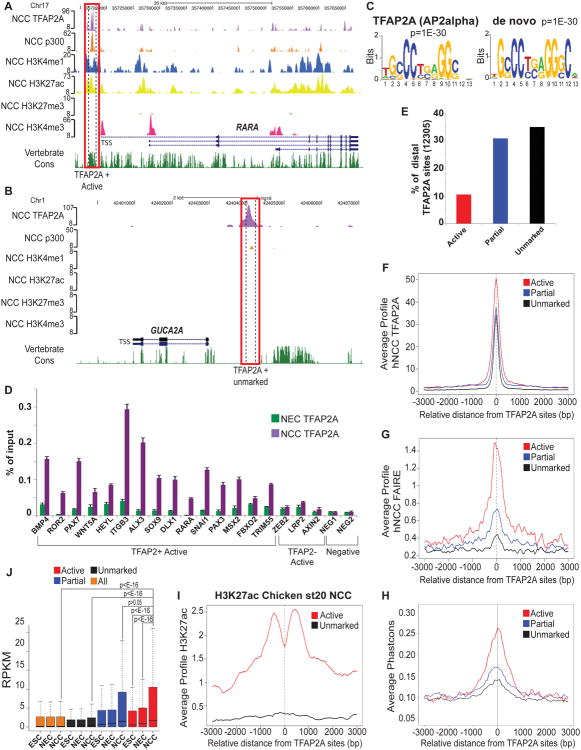
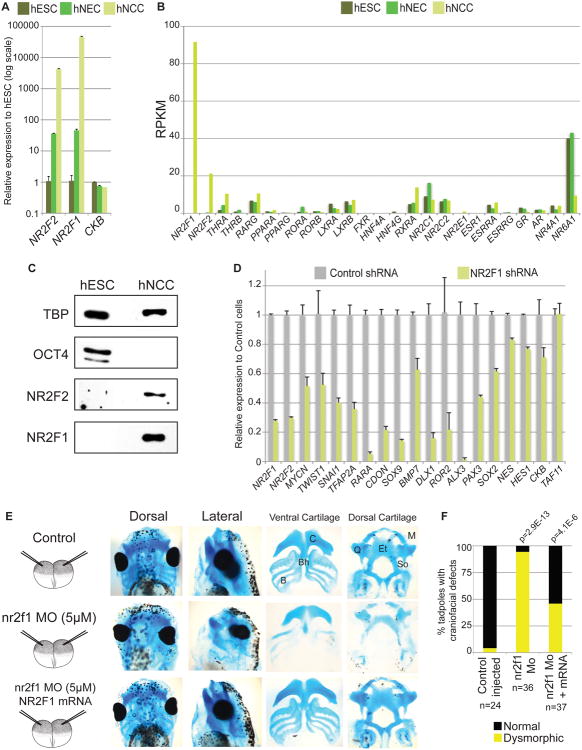
References
-
- Amiel J, Benko S, Gordon CT, Lyonnet S. Disruption of long-distance highly conserved noncoding elements in neurocristopathies. Ann N Y Acad Sci. 2010;1214:34–46. - PubMed
-
- Aybar MJ, Nieto MA, Mayor R. Snail precedes slug in the genetic cascade required for the specification and migration of the Xenopus neural crest. Development. 2003;130:483–494. - PubMed
-
- Bellmeyer A, Krase J, Lindgren J, LaBonne C. The protooncogene c-myc is an essential regulator of neural crest formation in xenopus. Dev Cell. 2003;4:827–839. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
