

The immune system and developmental programming of brain and behavior
- PMID: 22982535
- PMCID: PMC3484177
- DOI: 10.1016/j.yfrne.2012.08.006
The immune system and developmental programming of brain and behavior
Abstract
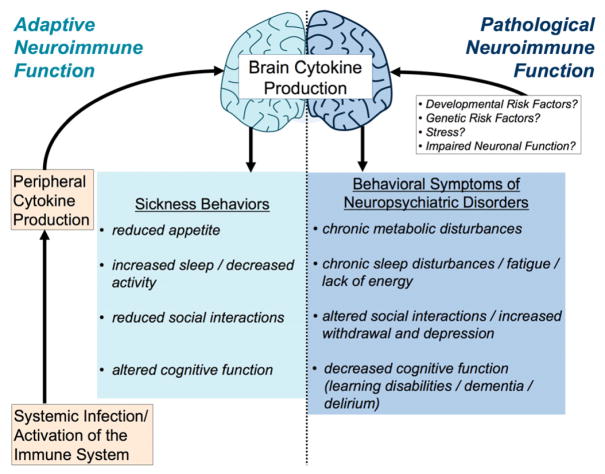
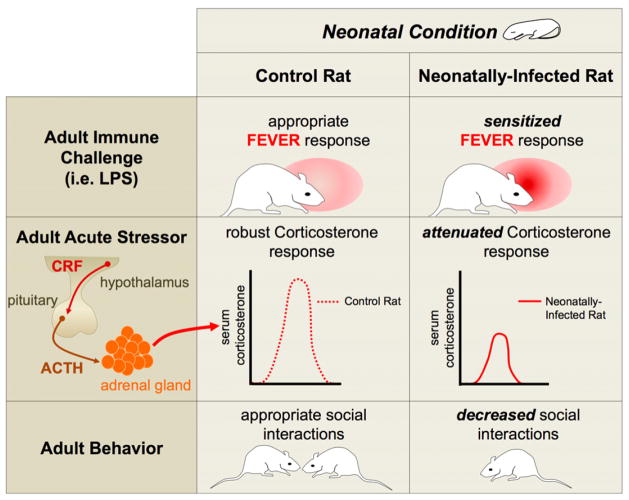
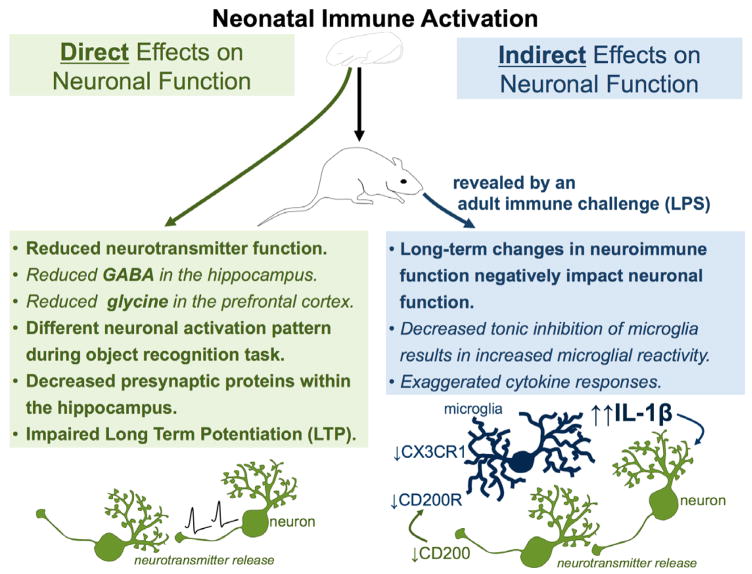
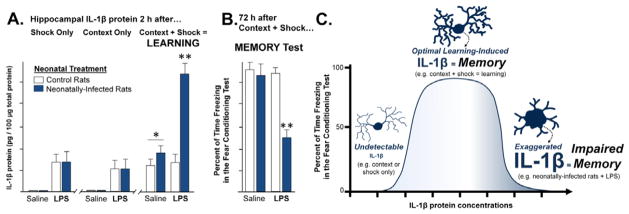
The brain, endocrine, and immune systems are inextricably linked. Immune molecules have a powerful impact on neuroendocrine function, including hormone-behavior interactions, during health as well as sickness. Similarly, alterations in hormones, such as during stress, can powerfully impact immune function or reactivity. These functional shifts are evolved, adaptive responses that organize changes in behavior and mobilize immune resources, but can also lead to pathology or exacerbate disease if prolonged or exaggerated. The developing brain in particular is exquisitely sensitive to both endogenous and exogenous signals, and increasing evidence suggests the immune system has a critical role in brain development and associated behavioral outcomes for the life of the individual. Indeed, there are associations between many neuropsychiatric disorders and immune dysfunction, with a distinct etiology in neurodevelopment. The goal of this review is to describe the important role of the immune system during brain development, and to discuss some of the many ways in which immune activation during early brain development can affect the later-life outcomes of neural function, immune function, mood and cognition.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
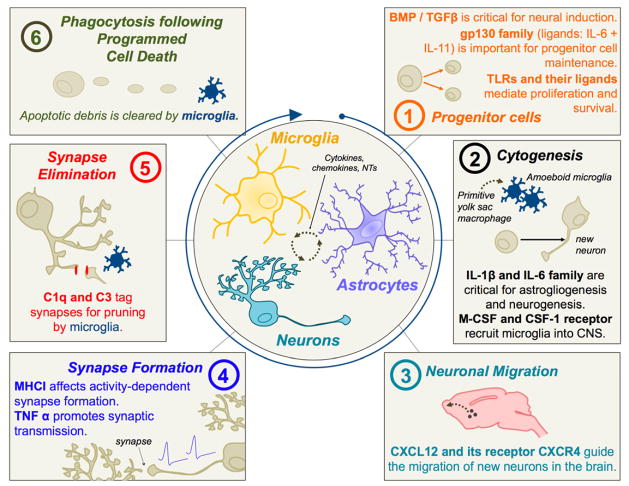
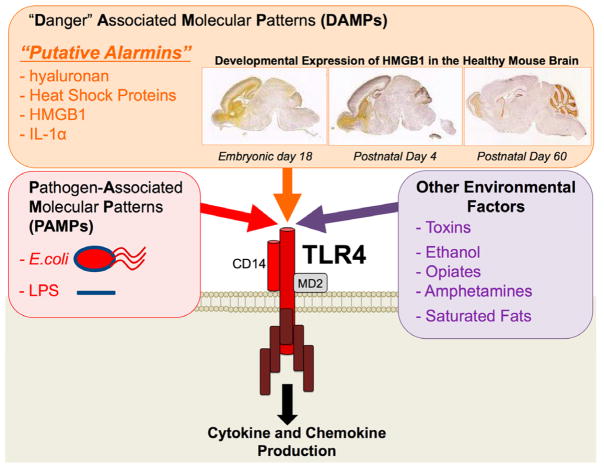
References
-
- Adams-Chapman I, Stoll BJ. Neonatal infection and long-term neurodevelopmental outcome in the preterm infant. Current opinion in infectious diseases. 2006;19:290–297. - PubMed
-
- Aiuti A, Webb IJ, Bleul C, Springer T, Gutierrez-Ramos JC. The chemokine SDF-1 is a chemoattractant for human CD34+ hematopoietic progenitor cells and provides a new mechanism to explain the mobilization of CD34+ progenitors to peripheral blood. The Journal of experimental medicine. 1997;185:111–120. - PMC - PubMed
-
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001;413:732–738. - PubMed
-
- Anderson KV, Jurgens G, Nusslein-Volhard C. Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell. 1985;42:779–789. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
