

Motor circuits are required to encode a sensory model for imitative learning
- PMID: 22983208
- PMCID: PMC3458123
- DOI: 10.1038/nn.3206
Motor circuits are required to encode a sensory model for imitative learning
Abstract
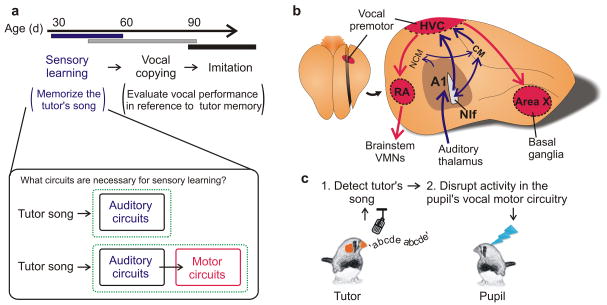
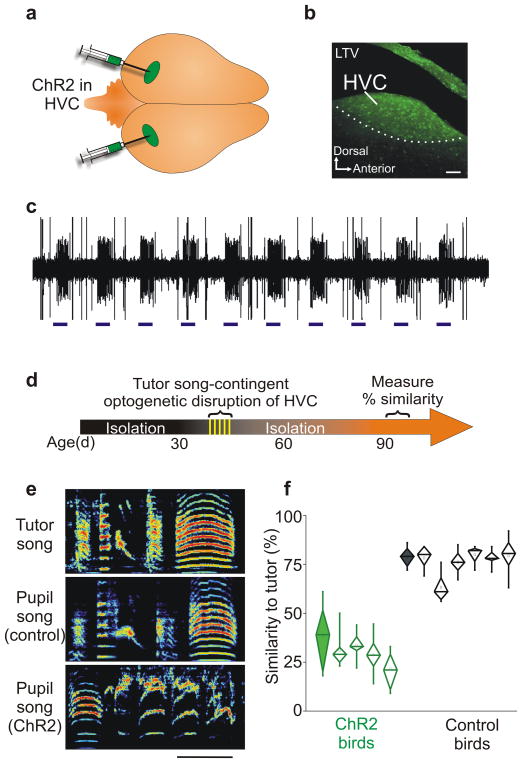
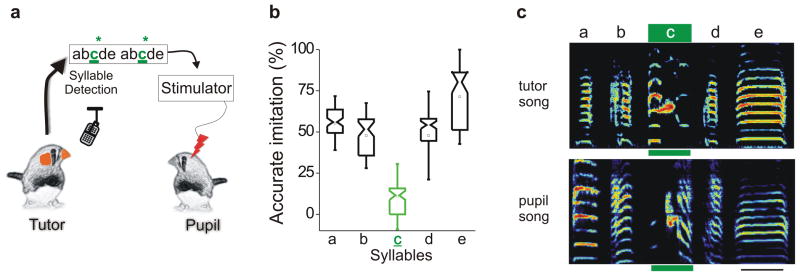
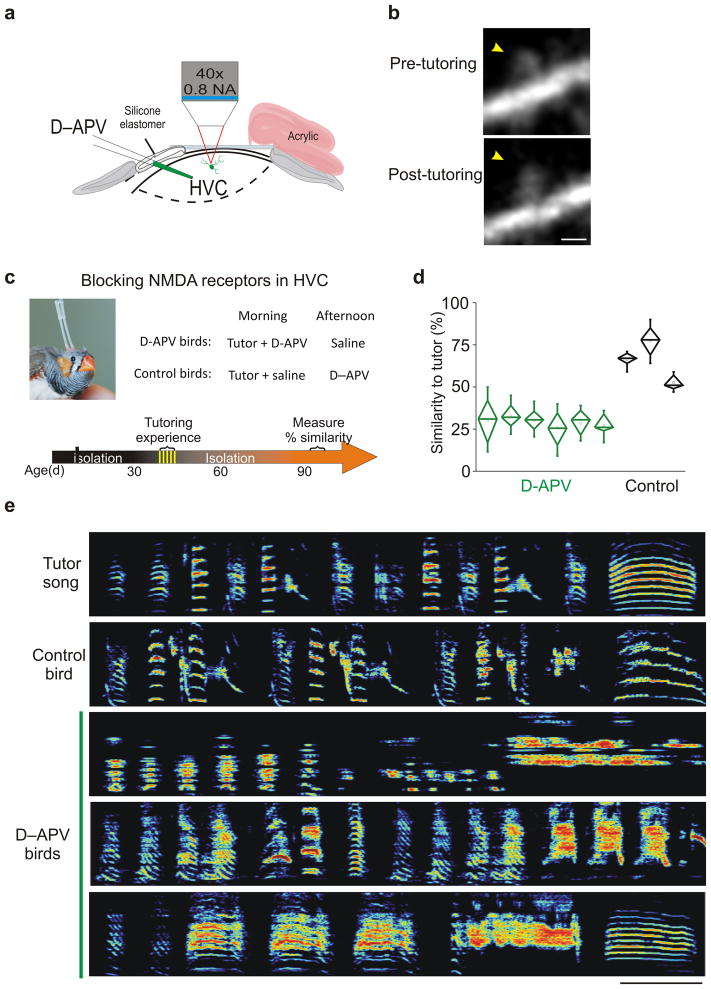
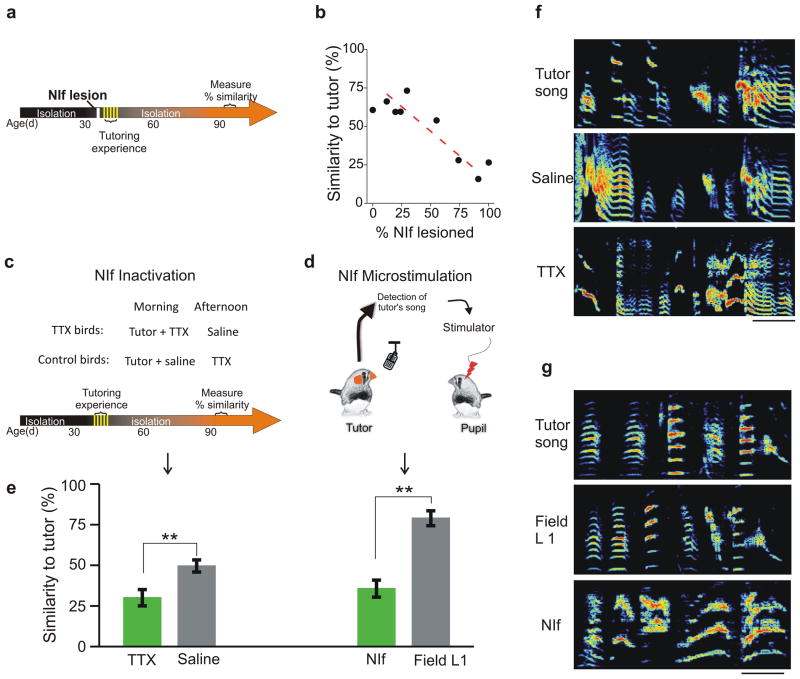
Premotor circuits help generate imitative behaviors and can be activated during observation of another animal's behavior, leading to speculation that these circuits participate in sensory learning that is important to imitation. Here we tested this idea by focally manipulating the brain activity of juvenile zebra finches, which learn to sing by memorizing and vocally copying the song of an adult tutor. Tutor song-contingent optogenetic or electrical disruption of neural activity in the pupil's song premotor nucleus HVC prevented song copying, indicating that a premotor structure important to the temporal control of birdsong also helps encode the tutor song. In vivo multiphoton imaging and neural manipulations delineated a pathway and a candidate synaptic mechanism through which tutor song information is encoded by premotor circuits. These findings provide evidence that premotor circuits help encode sensory information about the behavioral model before shaping and executing imitative behaviors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
A mesocortical dopamine circuit enables the cultural transmission of vocal behaviour.Nature. 2018 Nov;563(7729):117-120. doi: 10.1038/s41586-018-0636-7. Epub 2018 Oct 17. Nature. 2018. PMID: 30333629 Free PMC article.
-
Rapid spine stabilization and synaptic enhancement at the onset of behavioural learning.Nature. 2010 Feb 18;463(7283):948-52. doi: 10.1038/nature08759. Nature. 2010. PMID: 20164928 Free PMC article.
-
Experience-Dependent Intrinsic Plasticity During Auditory Learning.J Neurosci. 2019 Feb 13;39(7):1206-1221. doi: 10.1523/JNEUROSCI.1036-18.2018. Epub 2018 Dec 12. J Neurosci. 2019. PMID: 30541908 Free PMC article.
-
Neural mechanisms for learned birdsong.Learn Mem. 2009 Oct 22;16(11):655-69. doi: 10.1101/lm.1065209. Print 2009 Nov. Learn Mem. 2009. PMID: 19850665 Review.
-
Memory circuits for vocal imitation.Curr Opin Neurobiol. 2020 Feb;60:37-46. doi: 10.1016/j.conb.2019.11.002. Epub 2019 Dec 4. Curr Opin Neurobiol. 2020. PMID: 31810009 Free PMC article. Review.
Cited by
-
Expression of FoxP2 in the basal ganglia regulates vocal motor sequences in the adult songbird.Nat Commun. 2021 May 11;12(1):2617. doi: 10.1038/s41467-021-22918-2. Nat Commun. 2021. PMID: 33976169 Free PMC article.
-
A Hebbian learning rule gives rise to mirror neurons and links them to control theoretic inverse models.Front Neural Circuits. 2013 Jun 19;7:106. doi: 10.3389/fncir.2013.00106. eCollection 2013. Front Neural Circuits. 2013. PMID: 23801941 Free PMC article.
-
At the interface of the auditory and vocal motor systems: NIf and its role in vocal processing, production and learning.J Physiol Paris. 2013 Jun;107(3):178-92. doi: 10.1016/j.jphysparis.2013.04.001. Epub 2013 Apr 17. J Physiol Paris. 2013. PMID: 23603062 Free PMC article. Review.
-
Babbling opens the sensory phase for imitative vocal learning.Proc Natl Acad Sci U S A. 2024 Apr 30;121(18):e2312323121. doi: 10.1073/pnas.2312323121. Epub 2024 Apr 15. Proc Natl Acad Sci U S A. 2024. PMID: 38621117 Free PMC article.
-
A distributed neural network model for the distinct roles of medial and lateral HVC in zebra finch song production.J Neurophysiol. 2017 Aug 1;118(2):677-692. doi: 10.1152/jn.00917.2016. Epub 2017 Apr 5. J Neurophysiol. 2017. PMID: 28381490 Free PMC article.
References
-
- Ferrari PF, Gallese V, Rizzolatti G, Fogassi L. Mirror neurons responding to the observation of ingestive and communicative mouth actions in the monkey ventral premotor cortex. Eur J Neurosci. 2003;17:1703–1714. - PubMed
-
- Gallese V, Fadiga L, Fogassi L, Rizzolatti G. Action recognition in the premotor cortex. Brain. 1996;119 ( Pt 2):593–609. - PubMed
-
- Prather JF, Peters S, Nowicki S, Mooney R. Precise auditory-vocal mirroring in neurons for learned vocal communication. Nature. 2008;451:305–310. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
