

Novel system for efficient isolation of Clostridium double-crossover allelic exchange mutants enabling markerless chromosomal gene deletions and DNA integration
- PMID: 22983967
- PMCID: PMC3485963
- DOI: 10.1128/AEM.02214-12
Novel system for efficient isolation of Clostridium double-crossover allelic exchange mutants enabling markerless chromosomal gene deletions and DNA integration
Abstract
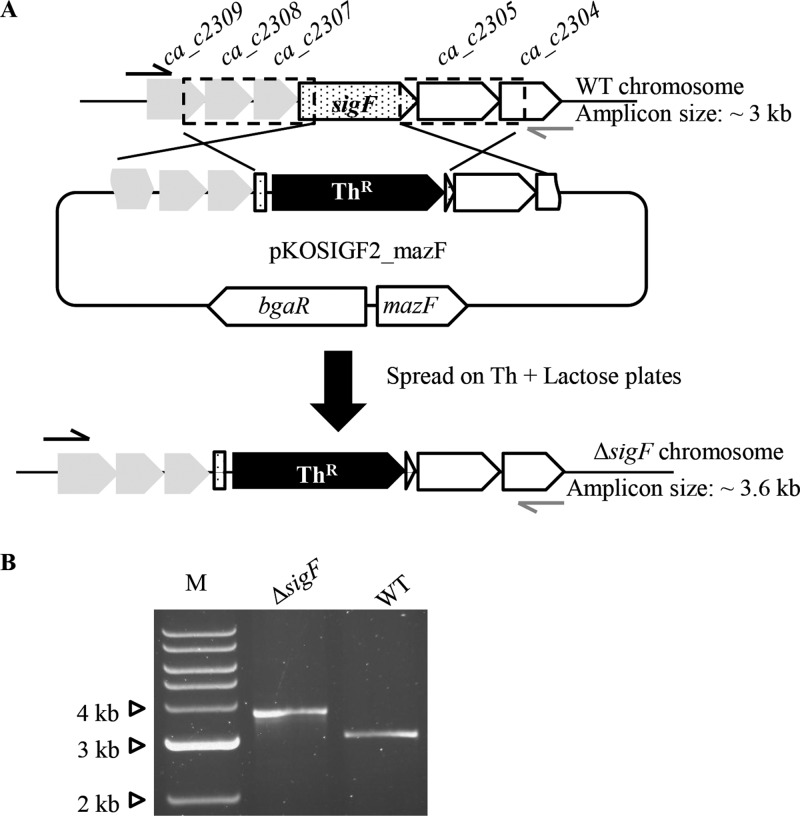
Isolation of Clostridium mutants based on gene replacement via allelic exchange remains a major limitation for this important genus. Use of a heterologous counterselection marker can facilitate the identification of the generally rare allelic exchange events. We report on the development of an inducible counterselection marker and describe its utility and broad potential in quickly and efficiently generating markerless DNA deletions and integrations at any genomic locus without the need for auxotrophic mutants or the use of the mobile group II introns. This system is based on a codon-optimized mazF toxin gene from Escherichia coli under the control of a lactose-inducible promoter from Clostridium perfringens. This system is potentially applicable to almost all members of the genus Clostridium due to their similarly low genomic GC content and comparable codon usage. We isolated all allelic-exchange-based gene deletions (ca_p0167, sigF, and sigK) or disruptions (ca_p0157 and sigF) we attempted and integrated a 3.6-kb heterologous DNA sequence (made up of a Clostridium ljungdahlii 2.1-kb formate dehydrogenase [fdh] gene plus a FLP recombination target [FRT]-flanked thiamphenicol resistance marker) into the Clostridium acetobutylicum chromosome. Furthermore, we report on the development of a plasmid system with inducible segregational instability, thus enabling efficient deployment of the FLP-FRT system to generate markerless deletion or integration mutants. This enabled expeditious deletion of the thiamphenicol resistance marker from the fdh integrant strain as well as the sigK deletion strain. More generally, our system can potentially be applied to other organisms with underdeveloped genetic tools.
Figures




References
-
- Awad MM, Bryant AE, Stevens DL, Rood JI. 1995. Virulence studies on chromosomal alpha-toxin and theta-toxin mutants constructed by allelic exchange provide genetic evidence for the essential role of alpha-toxin in Clostridium perfringens-mediated gas gangrene. Mol. Microbiol. 15: 191–202 - PubMed
-
- Boeke JD, Trueheart J, Natsoulis G, Fink GR. 1987. 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154: 164–175 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
