

A detailed, hierarchical study of Giardia lamblia's ventral disc reveals novel microtubule-associated protein complexes
- PMID: 22984443
- PMCID: PMC3439489
- DOI: 10.1371/journal.pone.0043783
A detailed, hierarchical study of Giardia lamblia's ventral disc reveals novel microtubule-associated protein complexes
Abstract
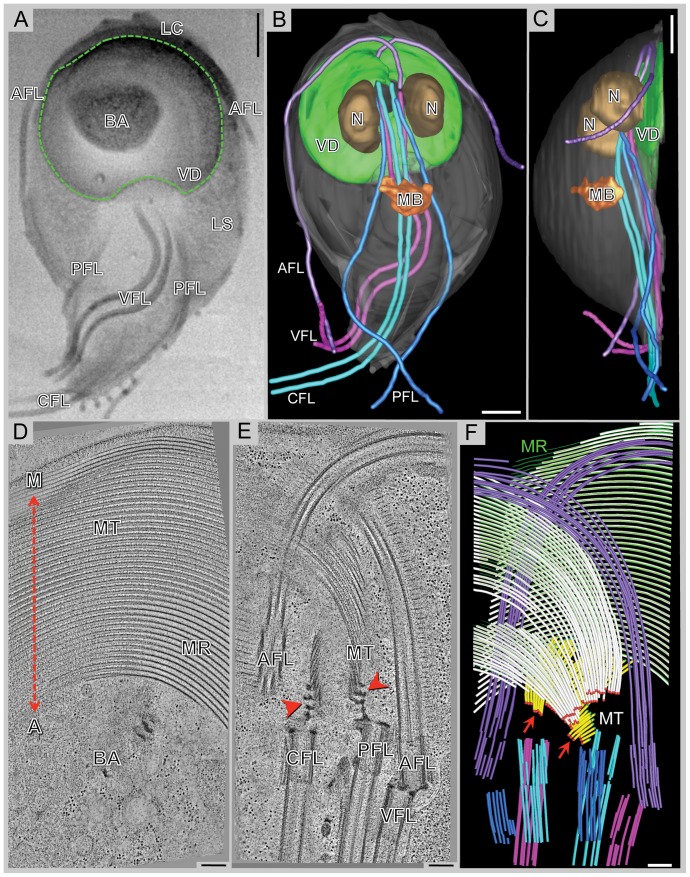
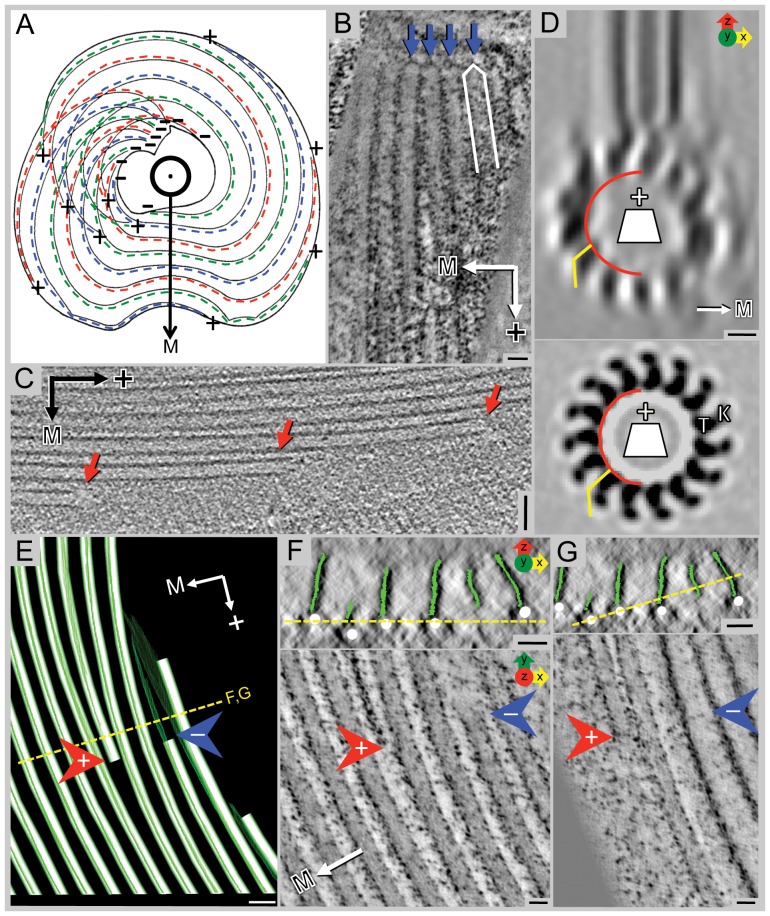
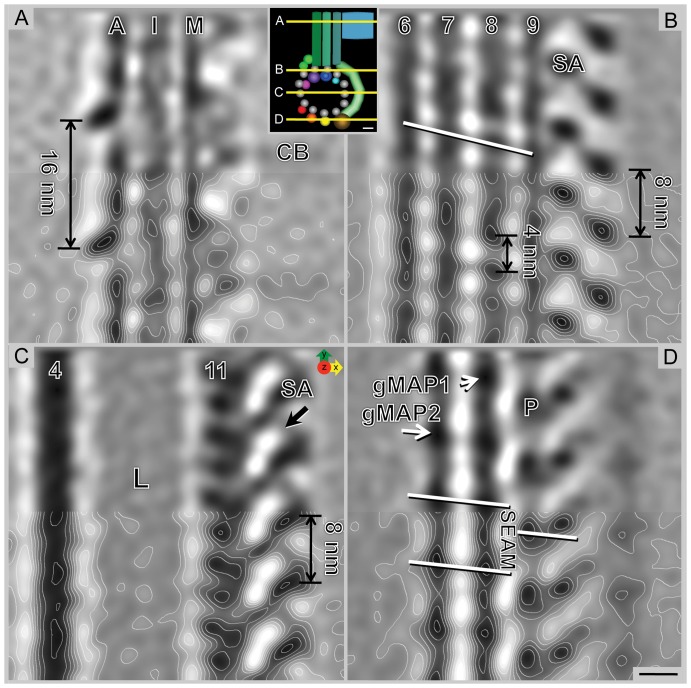
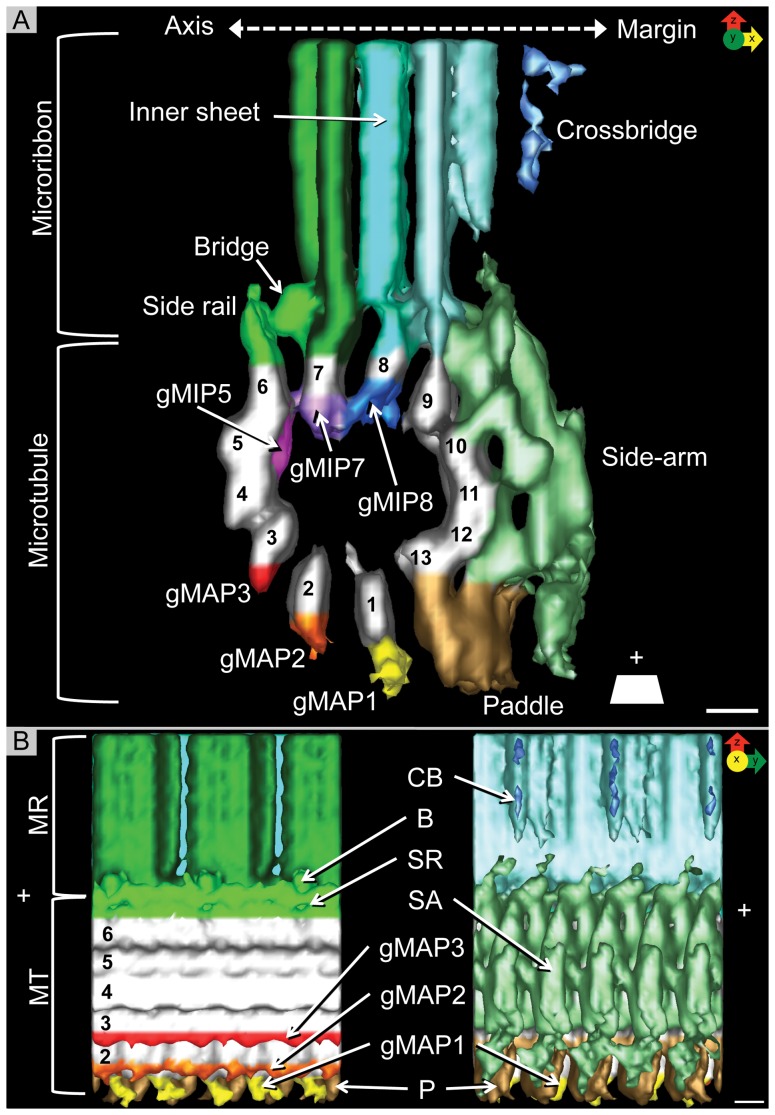
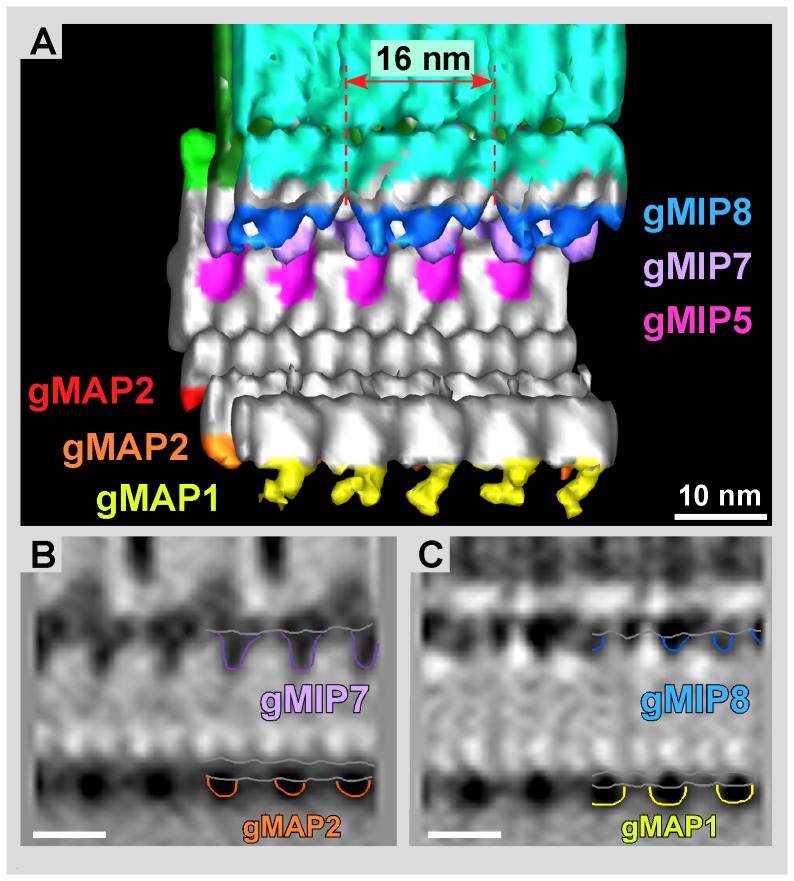
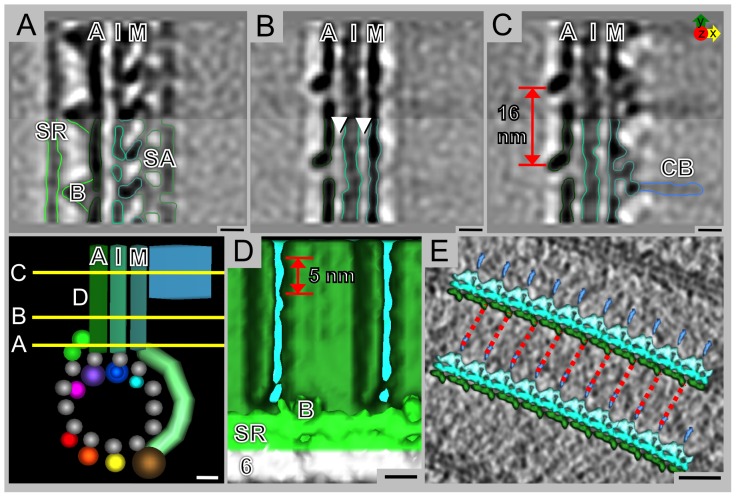
Giardia lamblia is a flagellated, unicellular parasite of mammals infecting over one billion people worldwide. Giardia's two-stage life cycle includes a motile trophozoite stage that colonizes the host small intestine and an infectious cyst form that can persist in the environment. Similar to many eukaryotic cells, Giardia contains several complex microtubule arrays that are involved in motility, chromosome segregation, organelle transport, maintenance of cell shape and transformation between the two life cycle stages. Giardia trophozoites also possess a unique spiral microtubule array, the ventral disc, made of approximately 50 parallel microtubules and associated microribbons, as well as a variety of associated proteins. The ventral disc maintains trophozoite attachment to the host intestinal epithelium. With the help of a combined SEM/microtome based slice and view method called 3View® (Gatan Inc., Pleasanton, CA), we present an entire trophozoite cell reconstruction and describe the arrangement of the major cytoskeletal elements. To aid in future analyses of disc-mediated attachment, we used electron-tomography of freeze-substituted, plastic-embedded trophozoites to explore the detailed architecture of ventral disc microtubules and their associated components. Lastly, we examined the disc microtubule array in three dimensions in unprecedented detail using cryo-electron tomography combined with internal sub-tomogram volume averaging of repetitive domains. We discovered details of protein complexes stabilizing microtubules by attachment to their inner and outer wall. A unique tri-laminar microribbon structure is attached vertically to the disc microtubules and is connected to neighboring microribbons via crossbridges. This work provides novel insight into the structure of the ventral disc microtubules, microribbons and associated proteins. Knowledge of the components comprising these structures and their three-dimensional organization is crucial toward understanding how attachment via the ventral disc occurs in vivo.
Conflict of interest statement
Figures

References
-
- Savioli L, Smith H, Thompson A (2006) Giardia and Cryptosporidium join the “Neglected Diseases Initiative”. Trends in parasitology 22: 203–208. - PubMed
-
- Thompson RC (2000) Giardiasis as a re-emerging infectious disease and its zoonotic potential. International journal for parasitology 30: 1259–1267. - PubMed
-
- Land KM, Johnson PJ (1999) Molecular basis of metronidazole resistance in pathogenic bacteria and protozoa. Drug resistance updates: reviews and commentaries in antimicrobial and anticancer chemotherapy 2: 289–294. - PubMed
