

Multiple roles for the actin cytoskeleton during regulated exocytosis
- PMID: 22986507
- PMCID: PMC4052552
- DOI: 10.1007/s00018-012-1156-5
Multiple roles for the actin cytoskeleton during regulated exocytosis
Abstract
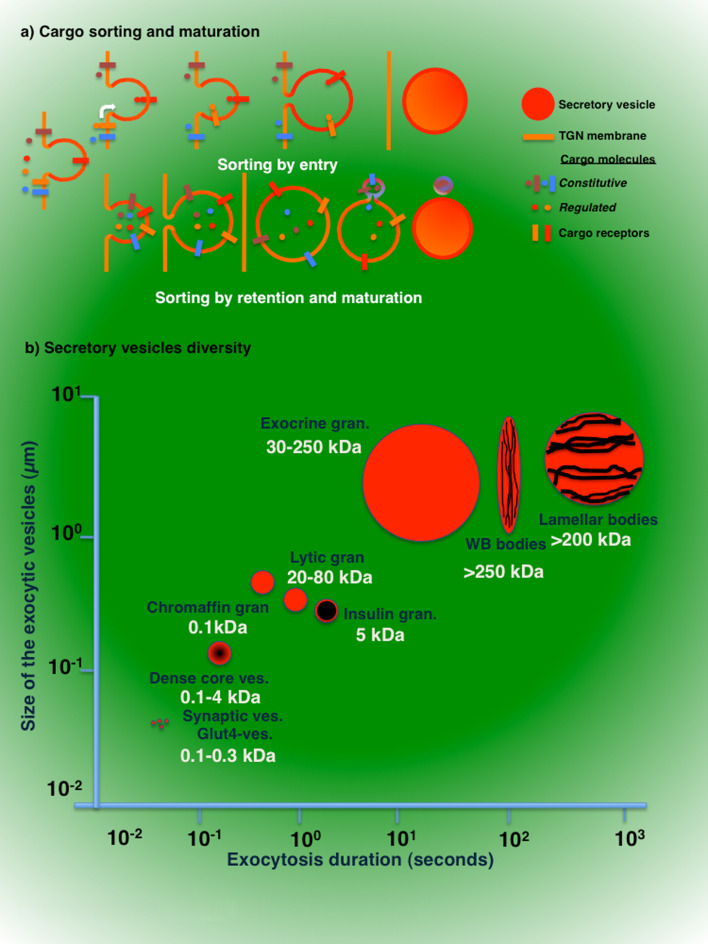
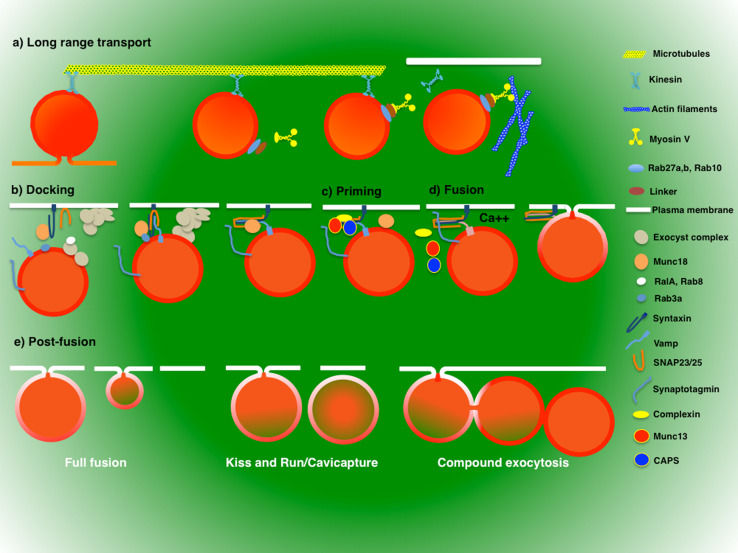
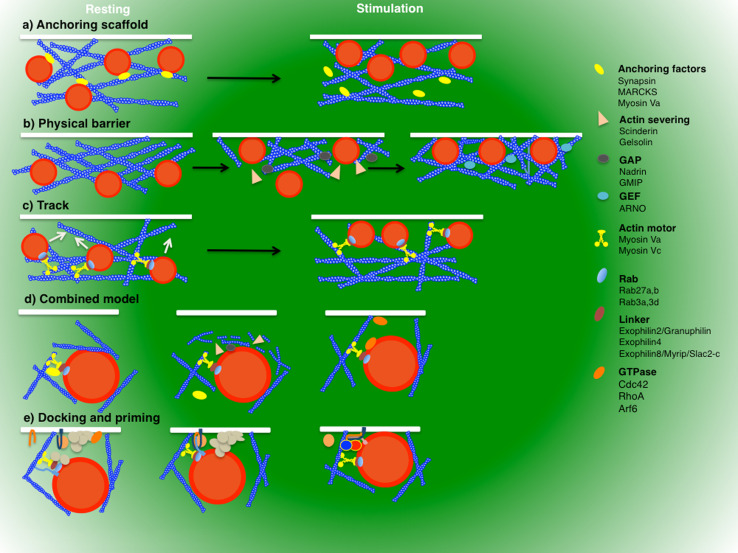
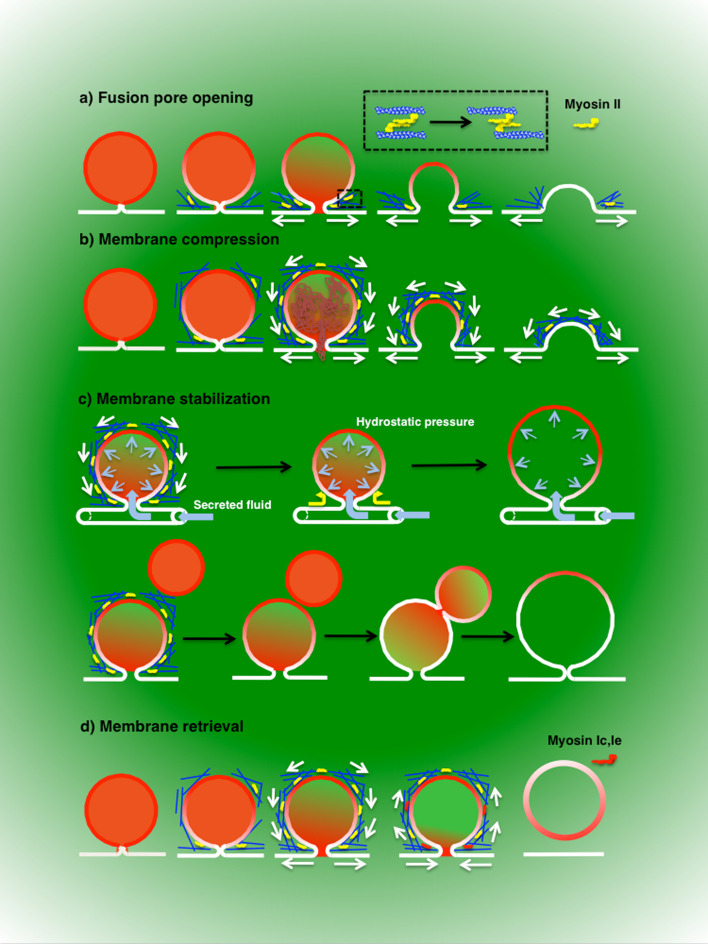
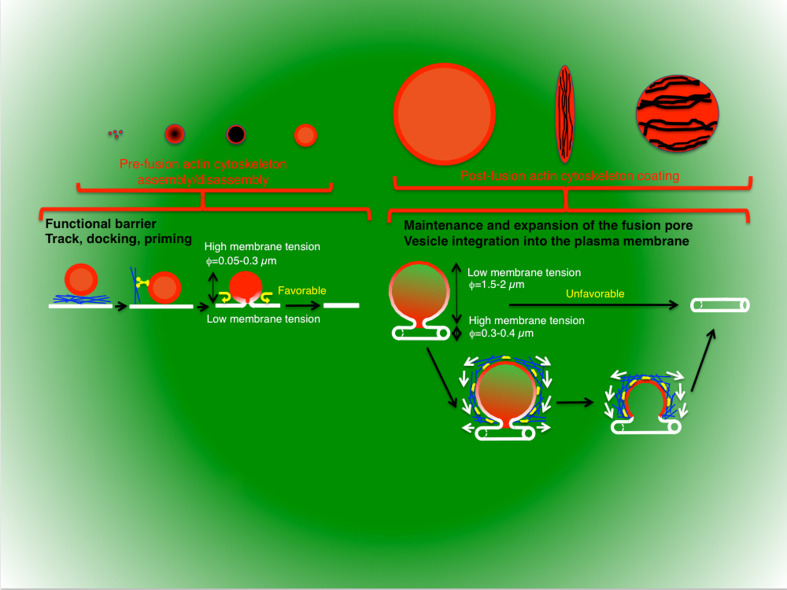
Regulated exocytosis is the main mechanism utilized by specialized secretory cells to deliver molecules to the cell surface by virtue of membranous containers (i.e., secretory vesicles). The process involves a series of highly coordinated and sequential steps, which include the biogenesis of the vesicles, their delivery to the cell periphery, their fusion with the plasma membrane, and the release of their content into the extracellular space. Each of these steps is regulated by the actin cytoskeleton. In this review, we summarize the current knowledge regarding the involvement of actin and its associated molecules during each of the exocytic steps in vertebrates, and suggest that the overall role of the actin cytoskeleton during regulated exocytosis is linked to the architecture and the physiology of the secretory cells under examination. Specifically, in neurons, neuroendocrine, endocrine, and hematopoietic cells, which contain small secretory vesicles that undergo rapid exocytosis (on the order of milliseconds), the actin cytoskeleton plays a role in pre-fusion events, where it acts primarily as a functional barrier and facilitates docking. In exocrine and other secretory cells, which contain large secretory vesicles that undergo slow exocytosis (seconds to minutes), the actin cytoskeleton plays a role in post-fusion events, where it regulates the dynamics of the fusion pore, facilitates the integration of the vesicles into the plasma membrane, provides structural support, and promotes the expulsion of large cargo molecules.
Figures
References
-
- Burgoyne RD, Morgan A. Secretory granule exocytosis. Physiol Rev. 2003;83(2):581–632. - PubMed
-
- De Matteis MA, Luini A. Exiting the Golgi complex. Nat Rev Mol Cell Biol. 2008;9(4):273–284. - PubMed
-
- Bossi G, Griffiths GM. CTL secretory lysosomes: biogenesis and secretion of a harmful organelle. Semin Immunol. 2005;17(1):87–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
