

Interaction of cyclin-dependent kinase 12/CrkRS with cyclin K1 is required for the phosphorylation of the C-terminal domain of RNA polymerase II
- PMID: 22988298
- PMCID: PMC3486194
- DOI: 10.1128/MCB.06267-11
Interaction of cyclin-dependent kinase 12/CrkRS with cyclin K1 is required for the phosphorylation of the C-terminal domain of RNA polymerase II
Abstract
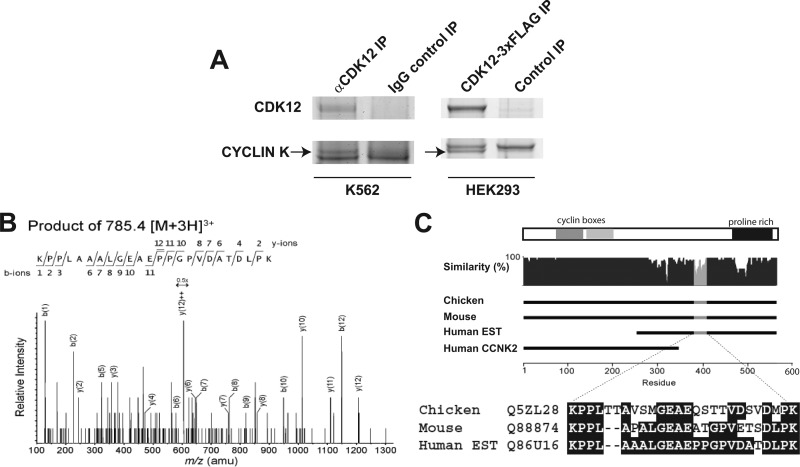
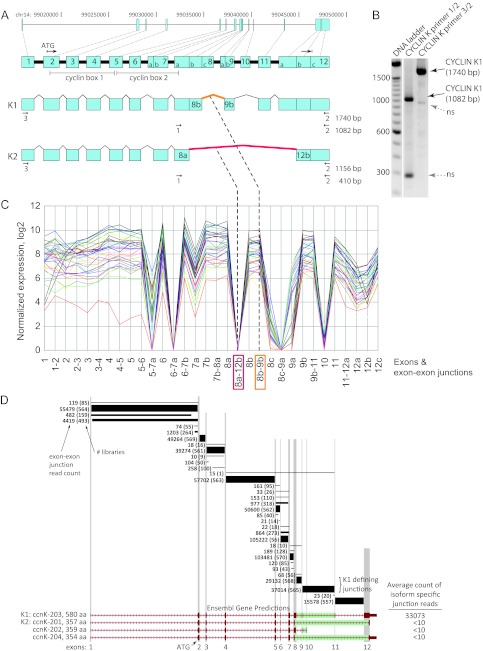
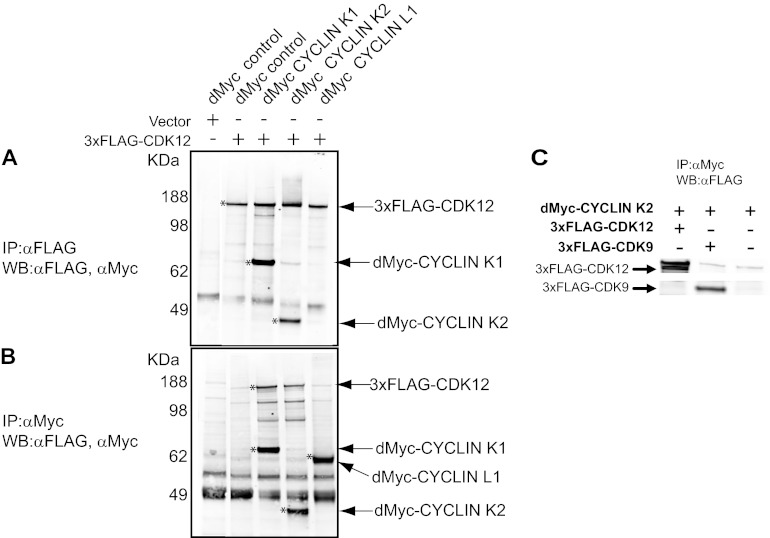
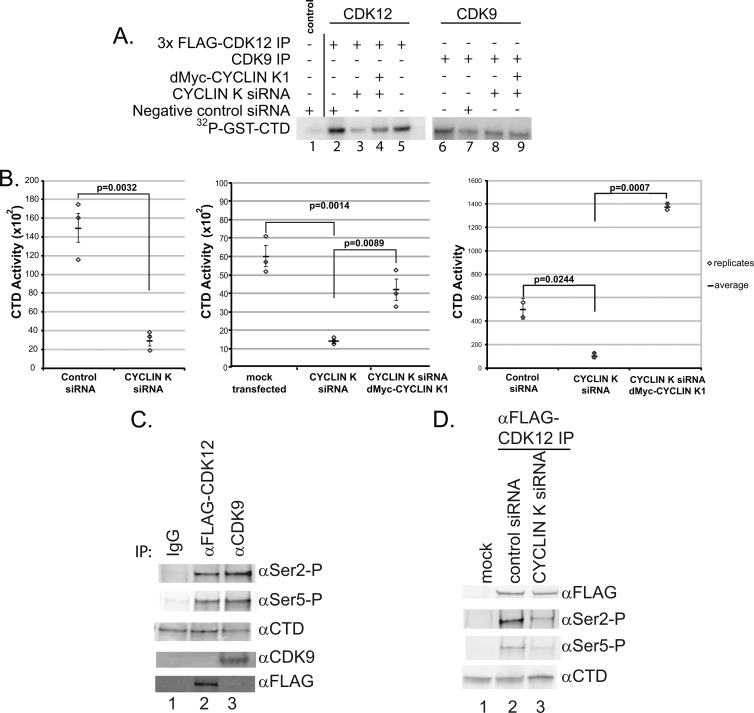
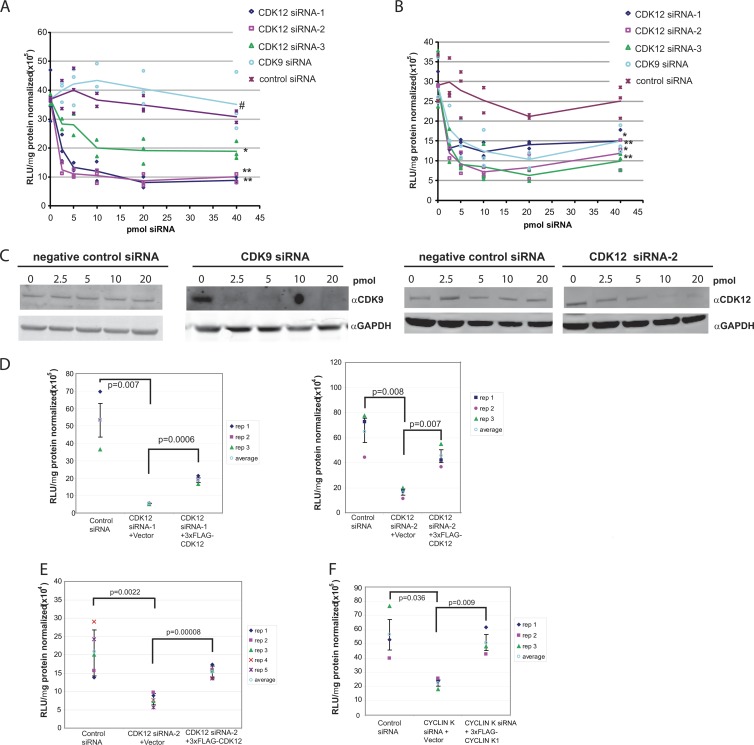
CrkRS (Cdc2-related kinase, Arg/Ser), or cyclin-dependent kinase 12 (CKD12), is a serine/threonine kinase believed to coordinate transcription and RNA splicing. While CDK12/CrkRS complexes were known to phosphorylate the C-terminal domain (CTD) of RNA polymerase II (RNA Pol II), the cyclin regulating this activity was not known. Using immunoprecipitation and mass spectrometry, we identified a 65-kDa isoform of cyclin K (cyclin K1) in endogenous CDK12/CrkRS protein complexes. We show that cyclin K1 complexes isolated from mammalian cells contain CDK12/CrkRS but do not contain CDK9, a presumed partner of cyclin K. Analysis of extensive RNA-Seq data shows that the 65-kDa cyclin K1 isoform is the predominantly expressed form across numerous tissue types. We also demonstrate that CDK12/CrkRS is dependent on cyclin K1 for its kinase activity and that small interfering RNA (siRNA) knockdown of CDK12/CrkRS or cyclin K1 has similar effects on the expression of a luciferase reporter gene. Our data suggest that cyclin K1 is the primary cyclin partner for CDK12/CrkRS and that cyclin K1 is required to activate CDK12/CrkRS to phosphorylate the CTD of RNA Pol II. These properties are consistent with a role of CDK12/CrkRS in regulating gene expression through phosphorylation of RNA Pol II.
Figures
References
-
- Ahn SH, Kim M, Buratowski S. 2004. Phosphorylation of serine 2 within the RNA polymerase II C-terminal domain couples transcription and 3′ end processing. Mol. Cell 13:67–76 - PubMed
-
- Balk SP, Knudsen KE. 2008. AR, the cell cycle, and prostate cancer. Nucl. Recept. Signal. 6:e001 doi:10.1621/nrs.06001 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous