

Prolonged FGF signaling is necessary for lung and liver induction in Xenopus
- PMID: 22988910
- PMCID: PMC3514138
- DOI: 10.1186/1471-213X-12-27
Prolonged FGF signaling is necessary for lung and liver induction in Xenopus
Abstract
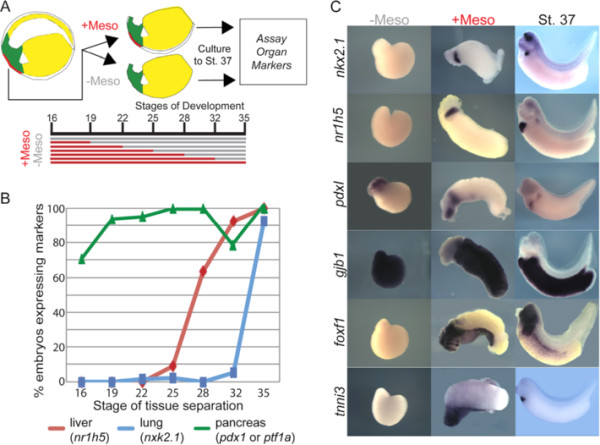
Background: FGF signaling plays numerous roles during organogenesis of the embryonic gut tube. Mouse explant studies suggest that different thresholds of FGF signaling from the cardiogenic mesoderm induce lung, liver, and pancreas lineages from the ventral foregut progenitor cells. The mechanisms that regulate FGF dose in vivo are unknown. Here we use Xenopus embryos to examine the hypothesis that a prolonged duration of FGF signaling from the mesoderm is required to induce foregut organs.
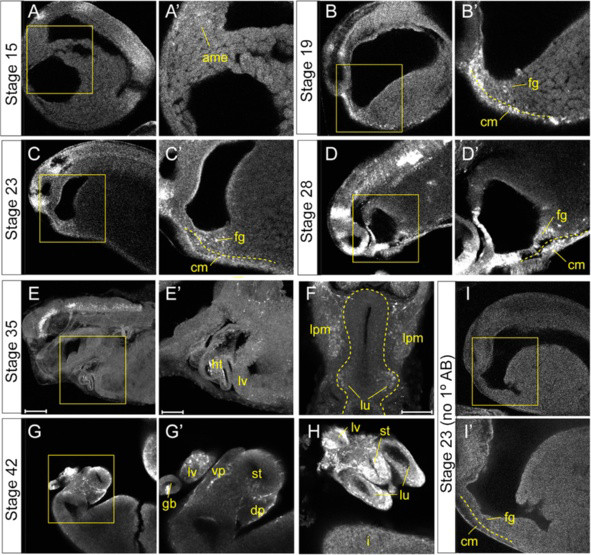
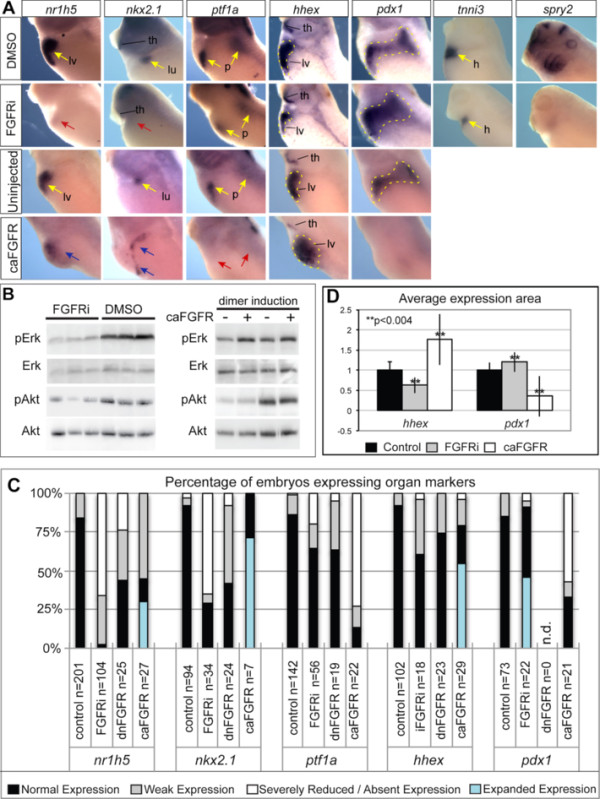
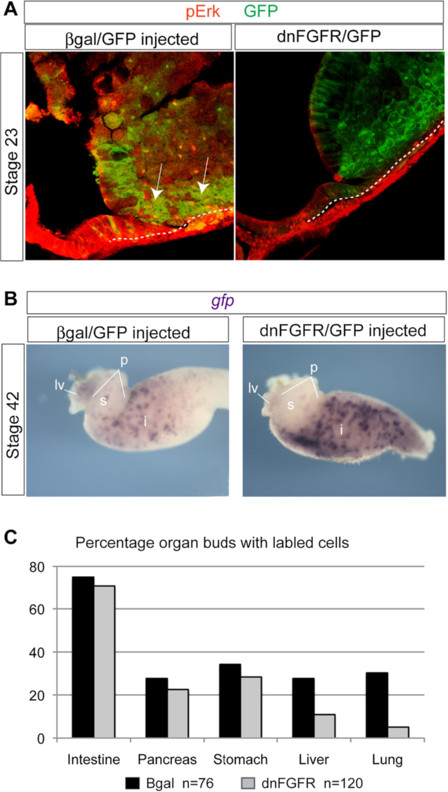
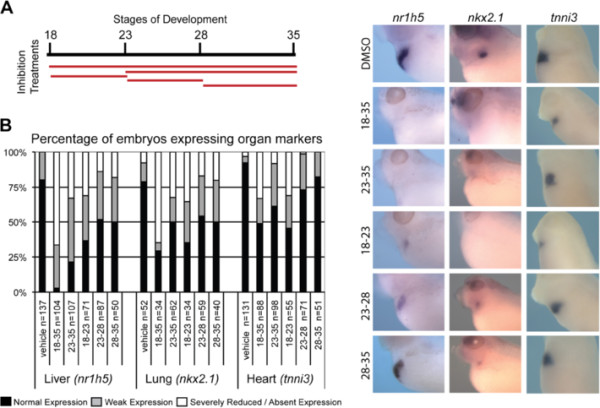
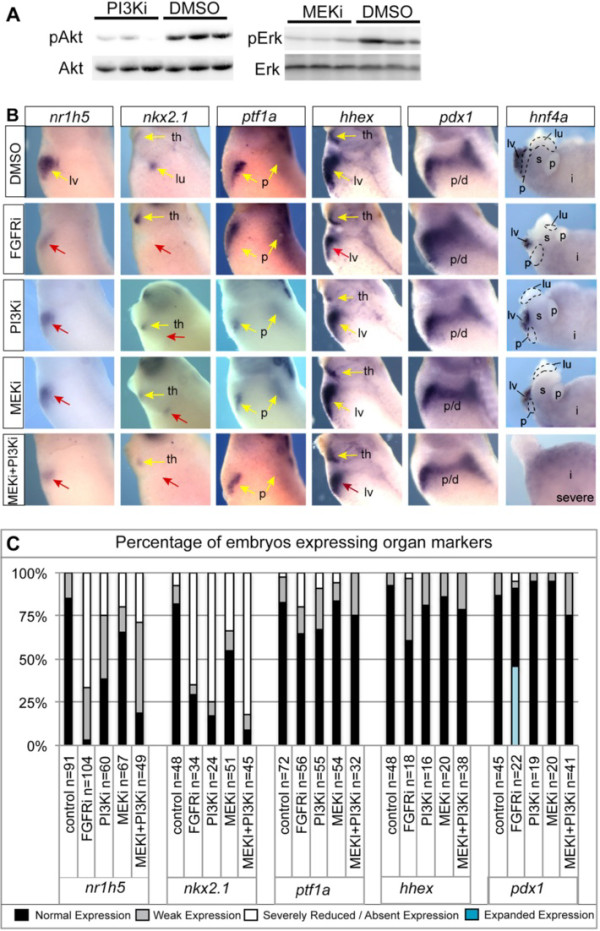
Results: We show that both mesoderm and FGF signaling are required for liver and lung development in Xenopus; formally demonstrating that this important step in organ induction is conserved with other vertebrate species. Prolonged contact with the mesoderm and persistent FGF signaling through both MEK and PI3K over an extended period of time are required for liver and lung specification. Inhibition of FGF signaling results in reduced liver and lung development, with a modest expansion of the pancreas/duodenum progenitor domain. Hyper-activation of FGF signaling has the opposite effect expanding liver and lung gene expression and repressing pancreatic markers. We show that FGF signaling is cell autonomously required in the endoderm and that a dominant negative FGF receptor decreases the ability of ventral foregut progenitor cells to contribute to the lung and liver buds.
Conclusions: These results suggest that the liver and lungs are specified at progressively later times in development requiring mesoderm contact for different lengths of time. Our data suggest that this is achieved at least in part through prolonged FGF signaling. In addition to providing a foundation for further mechanistic studies on foregut organogenesis using the experimental advantages of the Xenopus system, these data have implications for the directed differentiation of stem cells into foregut lineages.
Figures
Similar articles
-
Different thresholds of fibroblast growth factors pattern the ventral foregut into liver and lung.Development. 2005 Jan;132(1):35-47. doi: 10.1242/dev.01570. Epub 2004 Dec 2. Development. 2005. PMID: 15576401
-
FGF signaling is required for anterior but not posterior specification of the murine liver bud.Dev Dyn. 2015 Mar;244(3):431-43. doi: 10.1002/dvdy.24215. Epub 2014 Oct 23. Dev Dyn. 2015. PMID: 25302779 Free PMC article.
-
A Retinoic Acid-Hedgehog Cascade Coordinates Mesoderm-Inducing Signals and Endoderm Competence during Lung Specification.Cell Rep. 2016 Jun 28;16(1):66-78. doi: 10.1016/j.celrep.2016.05.060. Epub 2016 Jun 16. Cell Rep. 2016. PMID: 27320915 Free PMC article.
-
Signaling molecules that mediate the actions of FGF.Princess Takamatsu Symp. 1994;24:243-9. Princess Takamatsu Symp. 1994. PMID: 8983079 Review.
-
The role of mesodermal signals during liver organogenesis in zebrafish.Sci China Life Sci. 2010 Apr;53(4):455-61. doi: 10.1007/s11427-010-0078-4. Epub 2010 May 7. Sci China Life Sci. 2010. PMID: 20596911 Review.
Cited by
-
Syndecan4 coordinates Wnt/JNK and BMP signaling to regulate foregut progenitor development.Dev Biol. 2016 Aug 1;416(1):187-199. doi: 10.1016/j.ydbio.2016.05.025. Epub 2016 May 24. Dev Biol. 2016. PMID: 27235146 Free PMC article.
-
Wnt and FGF mediated epithelial-mesenchymal crosstalk during lung development.Dev Dyn. 2015 Mar;244(3):342-66. doi: 10.1002/dvdy.24234. Epub 2014 Dec 29. Dev Dyn. 2015. PMID: 25470458 Free PMC article. Review.
-
Temporal induction of the homeodomain transcription factor Nkx2-1 is sufficient to respecify foregut and hindgut endoderm to a pulmonary fate in Xenopus laevis.MicroPubl Biol. 2025 May 7;2025:10.17912/micropub.biology.001610. doi: 10.17912/micropub.biology.001610. eCollection 2025. MicroPubl Biol. 2025. PMID: 40406581 Free PMC article.
-
Identification of transcripts potentially involved in neural tube closure using RNA sequencing.Genesis. 2018 Mar;56(3):e23096. doi: 10.1002/dvg.23096. Epub 2018 Mar 10. Genesis. 2018. PMID: 29488319 Free PMC article.
-
Liver Specification in the Absence of Cardiac Differentiation Revealed by Differential Sensitivity to Wnt/β Catenin Pathway Activation.Front Physiol. 2019 Mar 5;10:155. doi: 10.3389/fphys.2019.00155. eCollection 2019. Front Physiol. 2019. PMID: 30890948 Free PMC article.
References
-
- Dessimoz J, Opoka R, Kordich JJ, Grapin-Botton A, Wells JM. FGF signaling is necessary for establishing gut tube domains along the anterior-posterior axis in vivo. Mech Dev. 2006;123:42–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
