

doi: 10.1093/nar/gks857.
Epub 2012 Sep 18.
The DUBm subunit Sgf11 is required for mRNA export and interacts with Cbp80 in Drosophila
Affiliations
- PMID: 22989713
- PMCID: PMC3510517
- DOI: 10.1093/nar/gks857
Item in Clipboard
The DUBm subunit Sgf11 is required for mRNA export and interacts with Cbp80 in Drosophila
Nucleic Acids Res.
2012 Nov.
Abstract
SAGA/TFTC is a histone acetyltransferase complex that has a second enzymatic activity because of the presence of a deubiquitination module (DUBm). Drosophila DUBm consists of Sgf11, ENY2 and Nonstop proteins. We show that Sgf11 has other DUBm-independent functions. It associates with Cbp80 component of the cap-binding complex and is thereby recruited onto growing messenger ribonucleic acid (mRNA); it also interacts with the AMEX mRNA export complex and is essential for hsp70 mRNA export, as well as for general mRNA export from the nucleus. Thus, Sgf11 functions as a component of both SAGA DUBm and the mRNA biogenesis machinery.
Figures
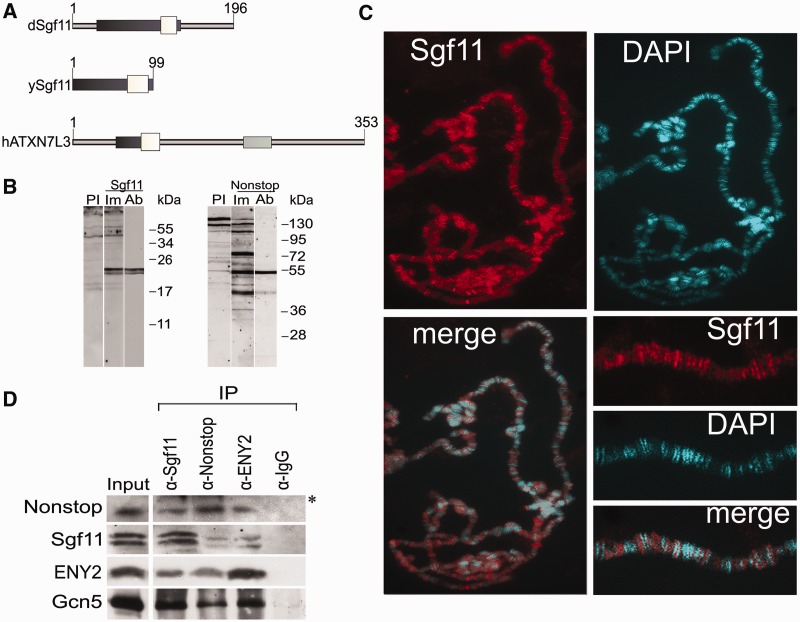
Nonstop, ENY2 and the moiety of Sgf11 form the Drosophila DUBm that interacts with SAGA. (A) Sgf11 structure and homology in different species (Drosophila, yeast and human). White boxes show homologous Sgf11-related domains; dark gray boxes, Sgf11 zinc finger-containing evolutionarily conserved regions and light gray boxes, the SCA7 domain, which is present only in the human protein. (B) Affinity-purified polyclonal antibodies raised in rabbits specifically recognize Sgf11 and Nonstop in nuclear extract from Drosophila embryos in western blot analysis. Lanes were stained with preimmune sera (PI), immune sera (Im) and affinity-purified polyclonal (Ab) antibodies are shown. (C) Sgf11 binds to numerous sites on polytene chromosomes from Drosophila salivary glands. Chromosomes stained with anti-Sgf11 antibodies and DAPI and merged images and scaled-up fragments of chromosomes are shown. (D) Sgf11 interacts with DUBm subunits and the Gcn5 component of SAGA in nuclear extract from Drosophila embryos in co-immunoprecipitation experiments (IP). Antibodies against Sgf11, Nonstop, ENY2 and Gcn5 or preimmune serum (PI) were used. Bands indicated with an asterisk correspond to antibodies.
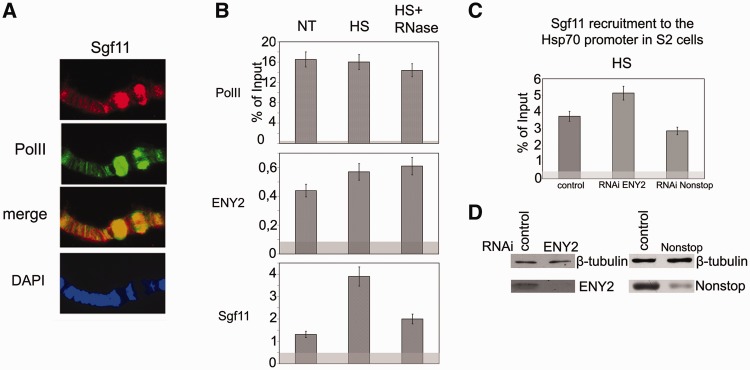
Sgf11 is present on the hsp70 promoter region. (A) Sgf11 is present in hsp70 puffs (87 A, 87B) on polytene chromosomes of Drosophila larvae under heat shock. Chromosomes were co-stained with antibodies against Sgf11, Pol II and DAPI. (B) Sgf11 occupancy of the hsp70 promoter increased after heat shock (HS) and significantly decreased after RNase treatment. The presence of Sgf11, Pol II and ENY2 was analyzed by ChIP before (NT) and after heat shock (HS). The results of ChIP are shown as a percentage of input. Here, and in Figures 2C and 6A–G, light gray shading indicates the background (baseline) level determined as the average of measurements in three noncoding sequences (see ‘Materials and Methods’ section). (C) Effects of ENY2 and Nonstop RNAi knockdown on Sgf11 recruitment on the hsp70 promoter under heat shock conditions (HS) according to the results of ChIP (shown as a percentage of input). Cells treated with GFP dsRNA were used as control. (D) The efficiency of ENY2 and Nonstop RNAi knockdown in experiments shown on Figure 3 C as verified by western blot analysis. Cells were treated with GFP dsRNA (control) or dsRNA corresponding to Sgf11 and Nonstop. Tubulin was used as loading control.
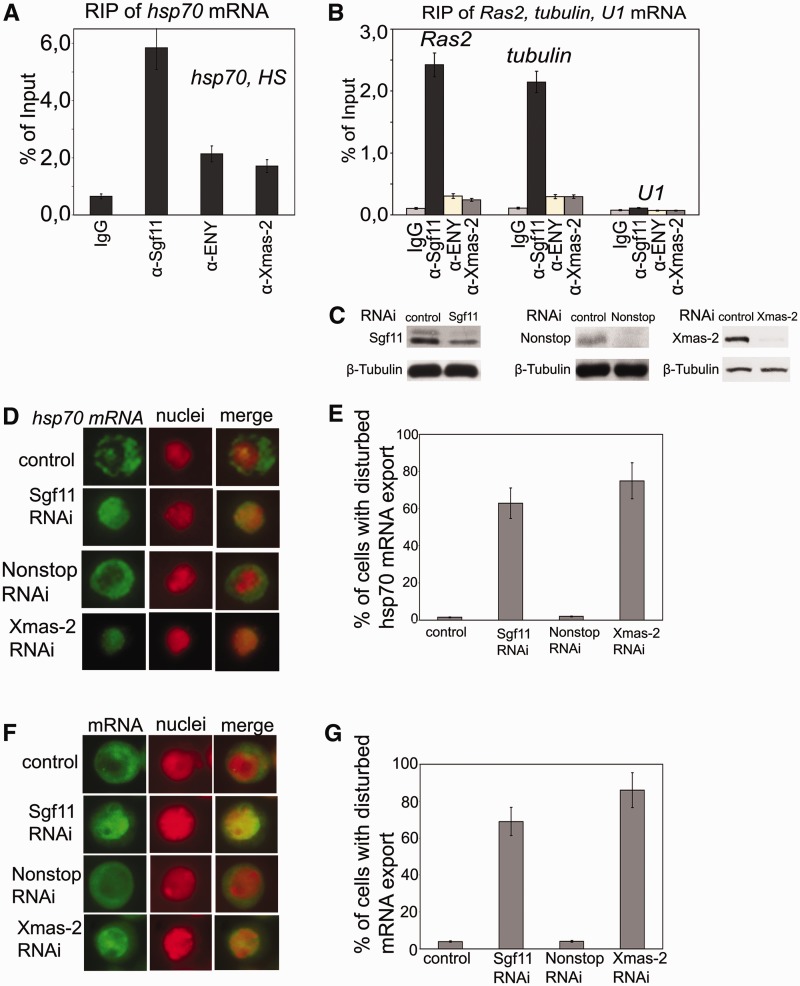
Sgf11 is associated with mRNAs of several genes, and its RNAi knockdown interferes with general mRNA export. (A) RIP experiments with hsp70 mRNA after heat shock were performed using antibodies against Sgf11 or components of the mRNA-interacting AMEX complex (ENY2, Xmas-2); nonimmune IgG was used as control. The results are shown as a percentage of input. (B) Sgf11 binds to mRNAs of Ras and tubulin genes under normal conditions. The U1 snRNA was used as a control. Antibodies used in RIP experiments were the same as in Figure 3 A. The results are shown as a percentage of input. (C) The level of Sgf11, Nonstop and Xmas-2 knockdown in experiments shown in Figures 3D–3G as estimated by western blot analysis in cells treated with GFP dsRNA (control) or dsRNA corresponding to Sgf11 and Nonstop. Tubulin was used as a loading control. (D) RNAi knockdown of Sgf11, but not Nonstop, interferes with hsp70 mRNA export after heat shock. Cells were treated with GFP dsRNA (control) or dsRNA corresponding to Sgf11 and Nonstop. Xmas-2 RNAi knockdown was performed as a positive control. Representative examples of the distribution of hsp70 mRNA (green staining) and cell nuclei (red staining) and corresponding merged images are shown for control cells and cells after Sgf11, Nonstop or Xmas-2 knockdown (magnification, ×1000). The hsp70 transcript was detected by RNA FISH using an Alexa 488-labeled probe; the nuclei were stained with DAPI. The images were recolored in Photoshop for better visualization. (E) Quantitative presentation of the results of experiments shown on Figure 3D. Bars show the percentage of cells with disturbed hsp70 mRNA nuclear export (about 200 cells per RNAi experiment were examined). (F) RNAi knockdown of Sgf11, but not Nonstop, interferes with general mRNA export. Cells were treated with GFP dsRNA (control) or dsRNA corresponding to Sgf11 and Nonstop. Xmas-2 RNAi knockdown was performed as a positive control. Representative examples of the distribution of mRNA (green staining) and cell nuclei (red staining) and corresponding merged images are shown for control cells and cells after Sgf11 or Nonstop knockdown (magnification, ×1000). RNA FISH was carried out using a Cy3-labeled oligo(dT) probe to identify poly(A) RNA. The nuclei were stained blue with DAPI. The images were recolored in Photoshop for better visualization. (G) Quantitative presentation of the results of experiments shown in Figure 3 F. Bars show the percentage of cells with disturbed hsp70 mRNA nuclear export (about 200 cells per RNAi experiment were examined).

Sgf11 is distributed between the nucleus and cytoplasm, co-localizes with NPC and interacts with AMEX. (A) Immunostaining of Drosophila S2 cells with antibodies against Sgf11 or Nonstop (red) and NPC (green) and merged images (magnification, ×1000). Panels c and d show cell fragments at higher magnification (×10000). (B) Sgf11 and Nonstop distribution in S2 cell nuclei and cytoplasmic extracts analyzed by western blotting. Equal amounts of extracts were loaded, and the blot was stained with antibodies against indicated proteins. Antibodies against TBP and β-tubulin were used to confirm that the cytoplasmic fraction was not contaminated with nuclear proteins, and vice versa. (C) Sgf11, but not Nonstop, interacts with Xmas-2, a component of the mRNA export complex AMEX, in co-immunoprecipitation from nuclear extract of Drosophila embryos. Antibodies against Xmas-2, Sgf11, Nonstop and IgG (control) were used. Western blot was stained with antibodies against Xmas-2. (D) Sgf11 RNAi knockdown does not affect the level of AMEX components in S2 cells. The presence of the indicated factors was analyzed in cells treated with control nonspecific dsRNA (GFP) or corresponding dsRNA. Tubulin was used as loading control. The levels of corresponding proteins in sham-treated cells (left lanes) or in cells after RNAi treatment (right lanes) are shown.
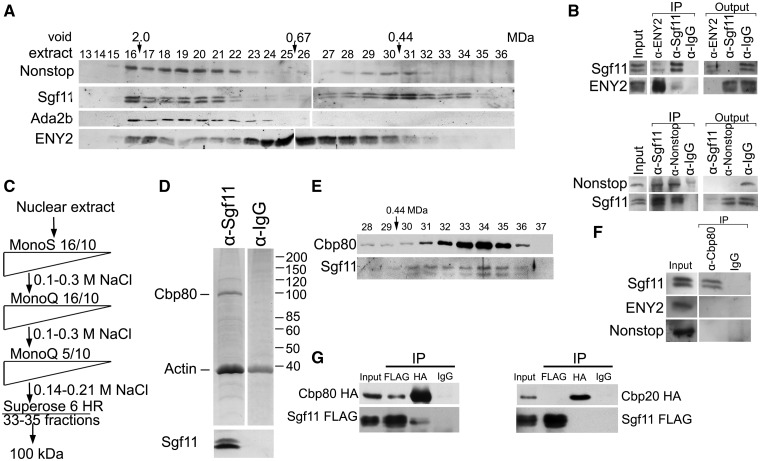
Sgf11 is present in several fractions of nuclear extract from Drosophila embryos and is associated with Cbp80 in a separate complex. (A) Sgf11 and Nonstop are present in low molecular weight fractions that do not contain SAGA complex. Drosophila embryonic extract treated with DNase I and RNase was fractionated on a Superose 6 gel filtration column. The fractions were analyzed for the presence of Sgf11, Nonstop, ENY2 and ADA2b (as a marker of SAGA distribution) by western blot analysis. Fraction numbers and the void volume are indicated. The distribution of ENY2, which has several characteristic peaks, testifies to the quality of fractionation. (B) Sgf11, Nonstop and ENY2 interact in low molecular weight fractions. Co-immunoprecipitation from the nuclear extract was performed with antibodies against Sgf11, Nonstop and ENY2 components of DUBm or with a preimmune serum (PI) in low molecular weight fractions 28–31. Equal amounts of the extract (Inp) and precipitated proteins (IP) were analyzed. (C) Scheme for purification of Sgf11-containing complexes. At each step, proteins were eluted with a NaCl gradient, and the fractions were analyzed for the presence of Sgf11 by western blotting. The peak Sgf11-containing fractions (the corresponding NaCl concentrations are indicated) were collected and loaded onto the next column. After Superose 6 fractionation, the material was loaded onto an immunosorbent with antibodies against Sgf11, washed and eluted with acid glycine. (D) Preparation of the Sgf11-containing complex purified from fractions 33–35 (∼100 kDa). Proteins eluted from the immunosorbent were resolved by 9% SDS-PAGE, stained with Coomassie and analyzed by mass spectrometry. The control immunoprecipitation of the same material with IgG is shown on the right. The bottom panels show western blot analysis of the Sgf11-containing preparation and the control immunoprecipitation for the presence of Sgf11. (E) The migration profiles of Cbp80 and Sgf11 on the Superose 6 column at the final purification step coincide with each other. (F) Co-immunoprecipitation experiments with nuclear extract of Drosophila embryos reveal no interactions between Cbp80 and ENY2 or Nonstop DUBm components. (G) Recombinant Sgf11 interact with Cbp80 but not with Cbp20. FLAG-tagged Sgf11 was co-expressed with HA-tagged Cbp80 or HA-tagged Cbp20 in transiently transfected S2 cells. Immunoprecipitation was performed with anti-FLAG or anti-HA antibodies or with IgG. Western blot was stained with anti-FLAG of anti-HA antibodies. About 10% of the input and 50 % of the precipitate were loaded onto the gel.
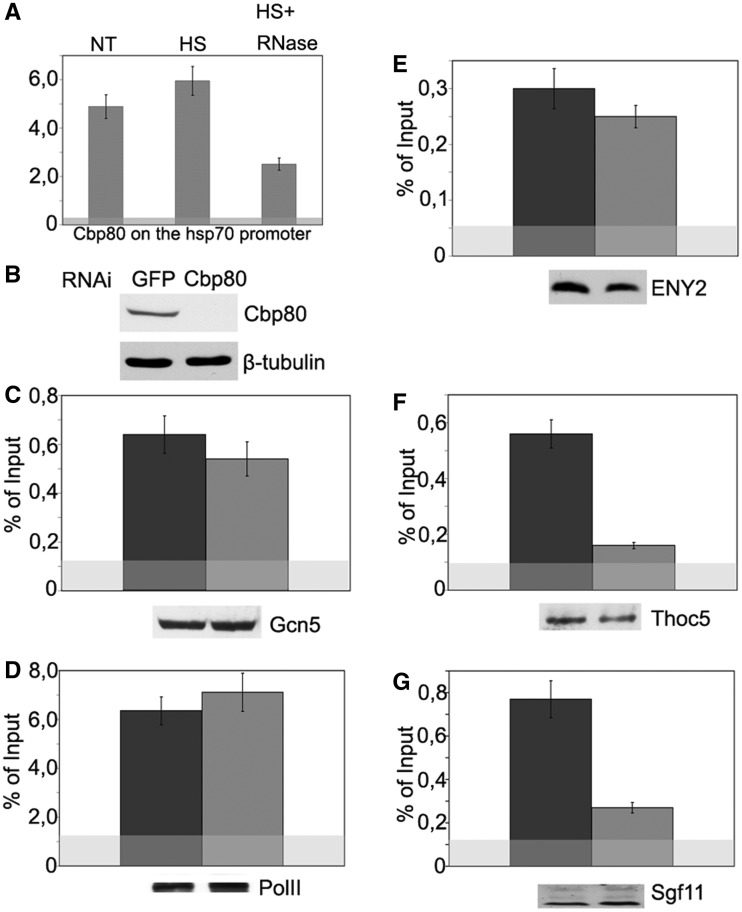
Sgf11 recruitment onto the hsp70 promoter region upon transcription activation depends on Cbp80. (A) Cbp80 occupancy of the hsp70 promoter as studied by ChIP. Chromatin was prepared before heat shock (NT), after heat shock (HS) or after heat shock with RNase treatment (HS+RNase). The results of ChIP are shown as a percentage of input. (B) Specificity of Cbp80 knockdown as tested by western blot analysis. The presence of Cbp80 was analyzed in cells treated with GFP dsRNA (control) or dsRNA. Tubulin was used as loading control. (C–G) The effect of Cbp80 RNAi knockdown on the recruitment of indicated proteins onto the hsp70 promoter after heat shock as studied by ChIP. Western blotting was used to confirm that Cbp80 knockdown did not affect the total level of test proteins in the cells (the results are shown below each figure).
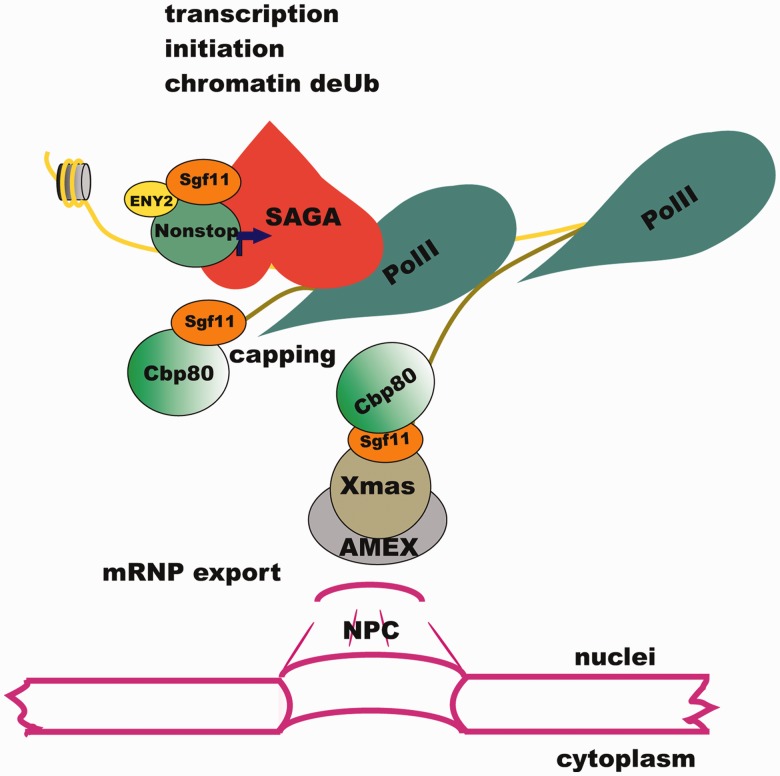
Sgf11 participates in different steps of gene expression in the nucleus. Sgf11 is localized on the promoter of the hsp70 gene as a component of DUBm of SAGA. Following transcription activation, Sgf11 interacts with the Cbp80 component of the CBC and associates with the growing transcript in proximity to chromatin. Sgf11 associates with mRNP, interacts with the AMEX mRNA export complex associated with NPC and is essential for mRNA nuclear export.
Similar articles
-
SAGA and a novel Drosophila export complex anchor efficient transcription and mRNA export to NPC.EMBO J. 2007 Dec 12;26(24):4956-65. doi: 10.1038/sj.emboj.7601901. Epub 2007 Nov 22. EMBO J. 2007. PMID: 18034162 Free PMC article.
-
Multifunctional factor ENY2 is associated with the THO complex and promotes its recruitment onto nascent mRNA.Genes Dev. 2010 Jan 1;24(1):86-96. doi: 10.1101/gad.550010. Genes Dev. 2010. PMID: 20048002 Free PMC article.
-
The Adaptor Protein ENY2 Is a Component of the Deubiquitination Module of the Arabidopsis SAGA Transcriptional Co-activator Complex but not of the TREX-2 Complex.J Mol Biol. 2018 May 11;430(10):1479-1494. doi: 10.1016/j.jmb.2018.03.018. Epub 2018 Mar 26. J Mol Biol. 2018. PMID: 29588169
-
[E(y)2, the novel component of SAGA/TFTC complex in eucaryotes participates in export of mRNP from nucleus and couples transcription with nuclear pore].Mol Biol (Mosk). 2009 Mar-Apr;43(2):253-63. Mol Biol (Mosk). 2009. PMID: 19425494 Review. Russian.
-
Dynamic modules of the coactivator SAGA in eukaryotic transcription.Exp Mol Med. 2020 Jul;52(7):991-1003. doi: 10.1038/s12276-020-0463-4. Epub 2020 Jul 3. Exp Mol Med. 2020. PMID: 32616828 Free PMC article. Review.
Cited by
-
Protein-coding circular RNA enhances antiviral immunity via JAK/STAT pathway in Drosophila.mBio. 2024 Sep 11;15(9):e0146924. doi: 10.1128/mbio.01469-24. Epub 2024 Aug 19. mBio. 2024. PMID: 39158293 Free PMC article.
-
Mammalian cells contain two functionally distinct PBAF complexes incorporating different isoforms of PHF10 signature subunit.Cell Cycle. 2014;13(12):1970-9. doi: 10.4161/cc.28922. Epub 2014 Apr 24. Cell Cycle. 2014. PMID: 24763304 Free PMC article.
-
Multifunctional ENY2 Protein Interacts with RNA Helicase MLE.Dokl Biochem Biophys. 2019 Nov;489(1):407-410. doi: 10.1134/S1607672919060140. Epub 2020 Mar 4. Dokl Biochem Biophys. 2019. PMID: 32130612
-
Cbp80 is needed for the expression of piRNA components and piRNAs.PLoS One. 2017 Jul 26;12(7):e0181743. doi: 10.1371/journal.pone.0181743. eCollection 2017. PLoS One. 2017. PMID: 28746365 Free PMC article.
-
Insulator protein Su(Hw) recruits SAGA and Brahma complexes and constitutes part of Origin Recognition Complex-binding sites in the Drosophila genome.Nucleic Acids Res. 2013 Jun;41(11):5717-30. doi: 10.1093/nar/gkt297. Epub 2013 Apr 22. Nucleic Acids Res. 2013. PMID: 23609538 Free PMC article.
References
-
- Balasubramanian R, Pray-Grant MG, Selleck W, Grant PA, Tan S. Role of the Ada2 and Ada3 transcriptional coactivators in histone acetylation. J. Biol. Chem. 2002;277:7989–7995. - PubMed
-
- Kuo MH, Brownell JE, Sobel RE, Ranalli TA, Cook RG, Edmondson DG, Roth SY, Allis CD. Transcription-linked acetylation by Gcn5p of histones H3 and H4 at specific lysines. Nature. 1996;383:269–272. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
