

Solution NMR structure of the Ca2+-bound N-terminal domain of CaBP7: a regulator of golgi trafficking
- PMID: 22989873
- PMCID: PMC3488092
- DOI: 10.1074/jbc.M112.402289
Solution NMR structure of the Ca2+-bound N-terminal domain of CaBP7: a regulator of golgi trafficking
Abstract
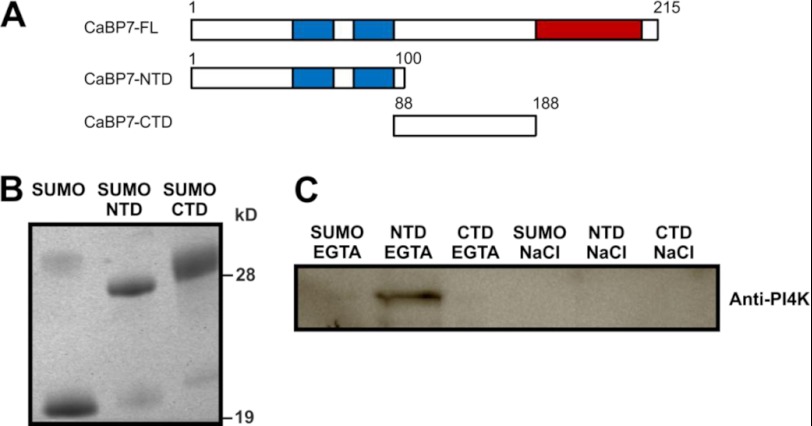
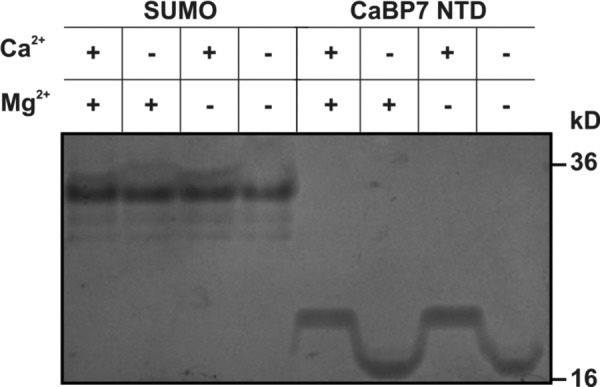
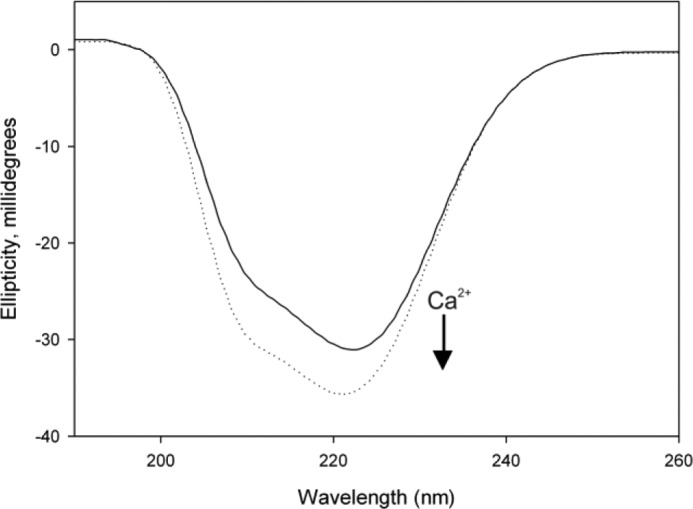
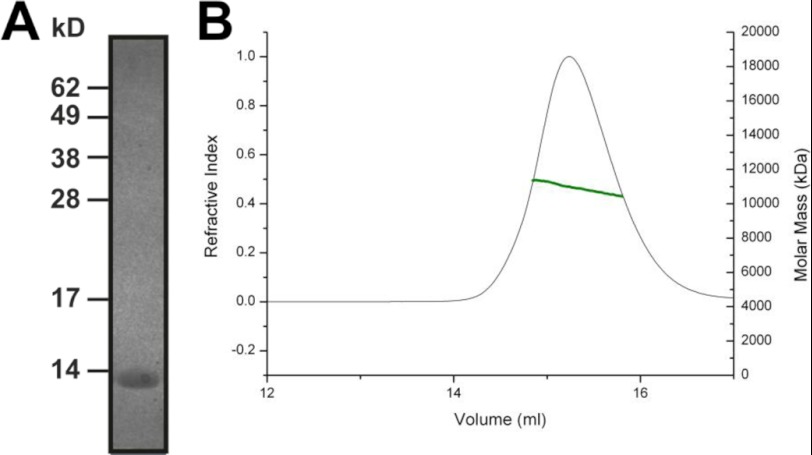
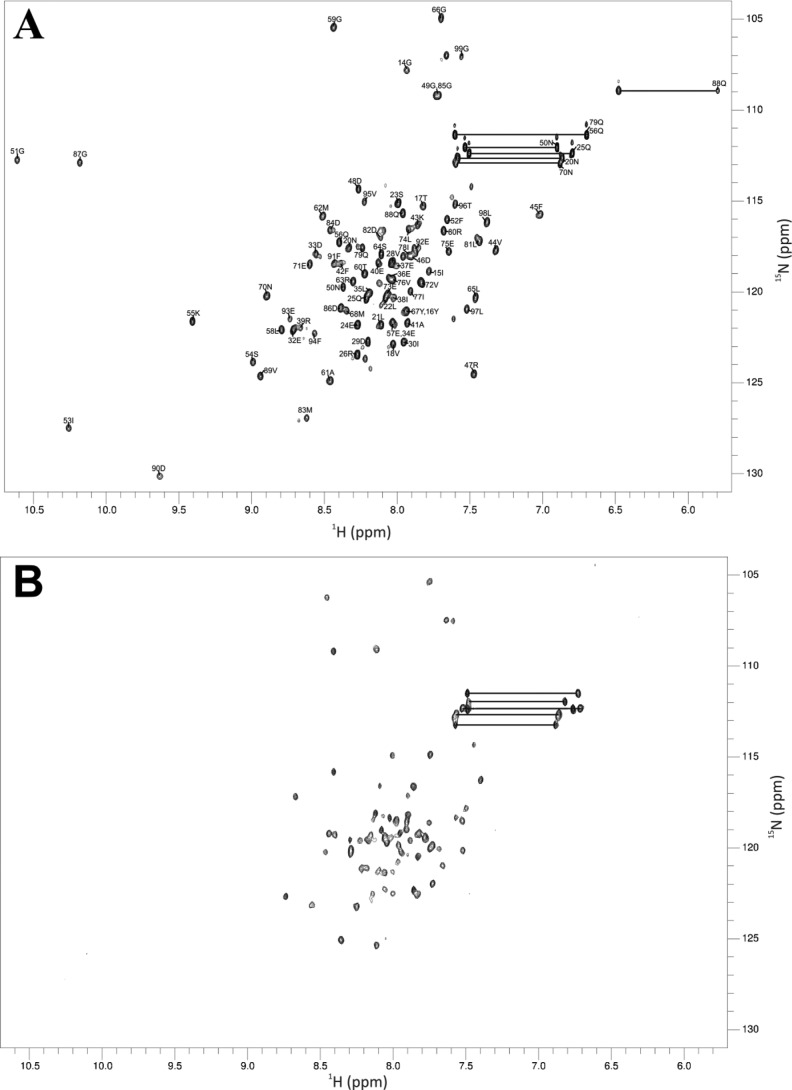
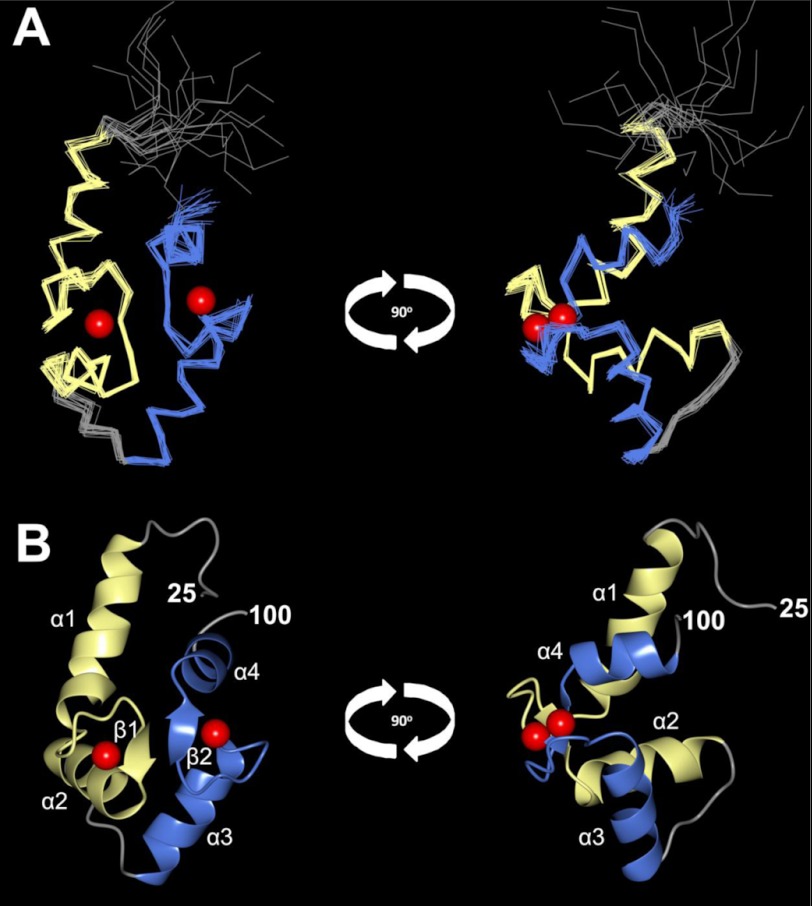
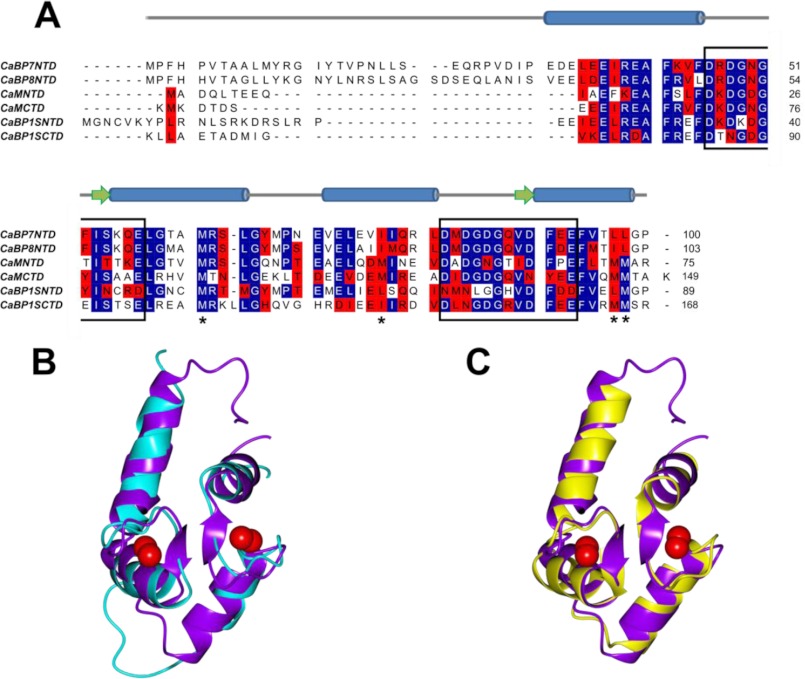
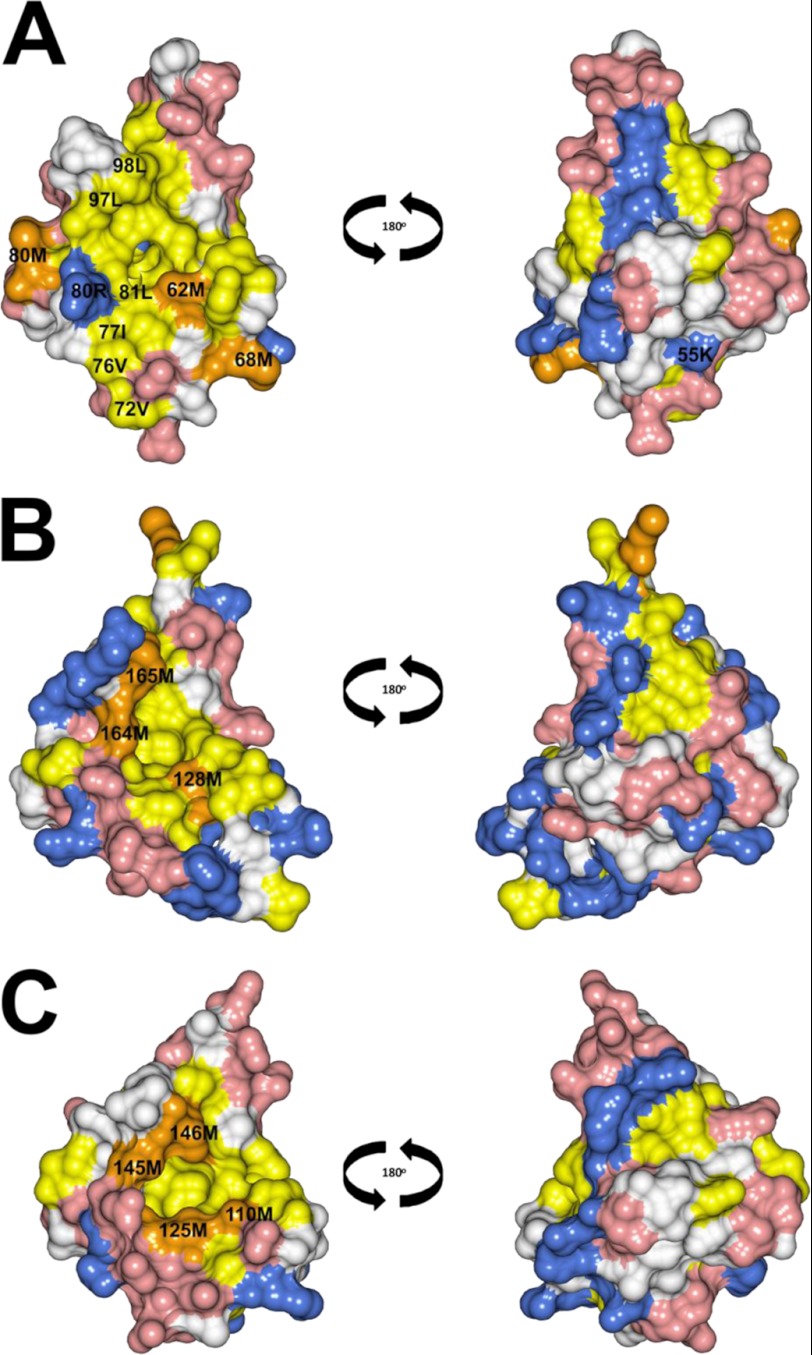
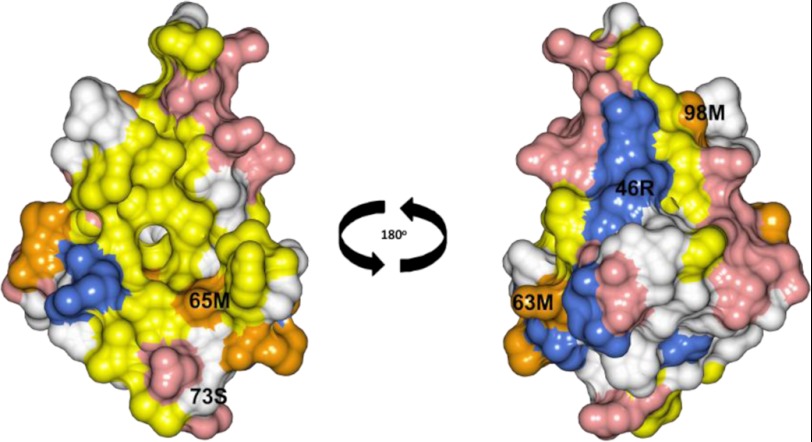
Calcium-binding protein 7 (CaBP7) is a member of the calmodulin (CaM) superfamily that harbors two high affinity EF-hand motifs and a C-terminal transmembrane domain. CaBP7 has been previously shown to interact with and modulate phosphatidylinositol 4-kinase III-β (PI4KIIIβ) activity in in vitro assays and affects vesicle transport in neurons when overexpressed. Here we show that the N-terminal domain (NTD) of CaBP7 is sufficient to mediate the interaction of CaBP7 with PI4KIIIβ. CaBP7 NTD encompasses the two high affinity Ca(2+) binding sites, and structural characterization through multiangle light scattering, circular dichroism, and NMR reveals unique properties for this domain. CaBP7 NTD binds specifically to Ca(2+) but not Mg(2+) and undergoes significant conformational changes in both secondary and tertiary structure upon Ca(2+) binding. The Ca(2+)-bound form of CaBP7 NTD is monomeric and exhibits an open conformation similar to that of CaM. Ca(2+)-bound CaBP7 NTD has a solvent-exposed hydrophobic surface that is more expansive than observed in CaM or CaBP1. Within this hydrophobic pocket, there is a significant reduction in the number of methionine residues that are conserved in CaM and CaBP1 and shown to be important for target recognition. In CaBP7 NTD, these residues are replaced with isoleucine and leucine residues with branched side chains that are intrinsically more rigid than the flexible methionine side chain. We propose that these differences in surface hydrophobicity, charge, and methionine content may be important in determining highly specific interactions of CaBP7 with target proteins, such as PI4KIIIβ.
Figures
References
-
- Seidenbecher C. I., Langnaese K., Sanmartí-Vila L., Boeckers T. M., Smalla K. H., Sabel B. A., Garner C. C., Gundelfinger E. D., Kreutz M. R. (1998) Caldendrin, a novel neuronal calcium-binding protein confined to the somato-dendritic compartment. J. Biol. Chem. 273, 21324–21331 - PubMed
-
- Wu Y. Q., Lin X., Liu C. M., Jamrich M., Shaffer L. G. (2001) Identification of a human brain-specific gene, calneuron 1, a new member of the calmodulin superfamily. Mol. Genet. Metab. 72, 343–350 - PubMed
-
- Mikhaylova M., Sharma Y., Reissner C., Nagel F., Aravind P., Rajini B., Smalla K. H., Gundelfinger E. D., Kreutz M. R. (2006) Neuronal Ca2+ signaling via caldendrin and calneurons. Biochim. Biophys. Acta 1763, 1229–1237 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
