

Minihepcidins prevent iron overload in a hepcidin-deficient mouse model of severe hemochromatosis
- PMID: 22990014
- PMCID: PMC3488893
- DOI: 10.1182/blood-2012-07-440743
Minihepcidins prevent iron overload in a hepcidin-deficient mouse model of severe hemochromatosis
Abstract
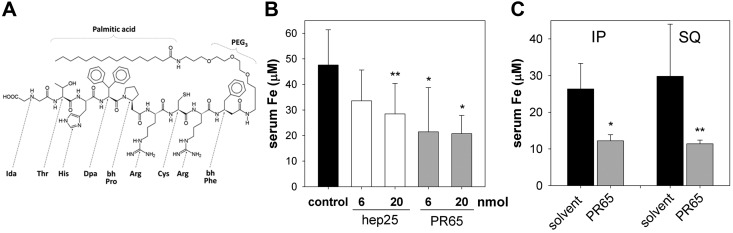
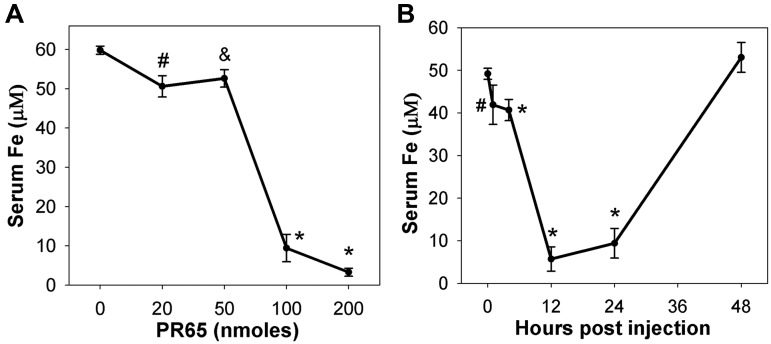
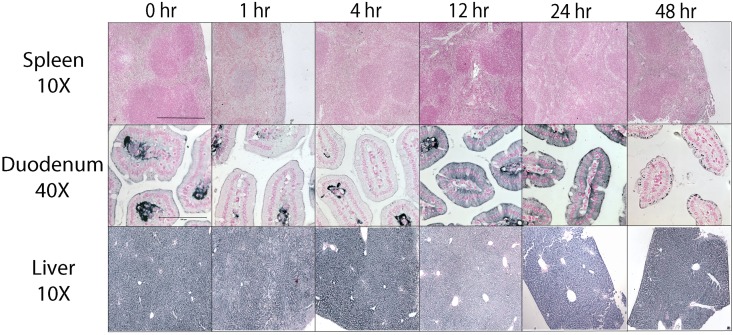
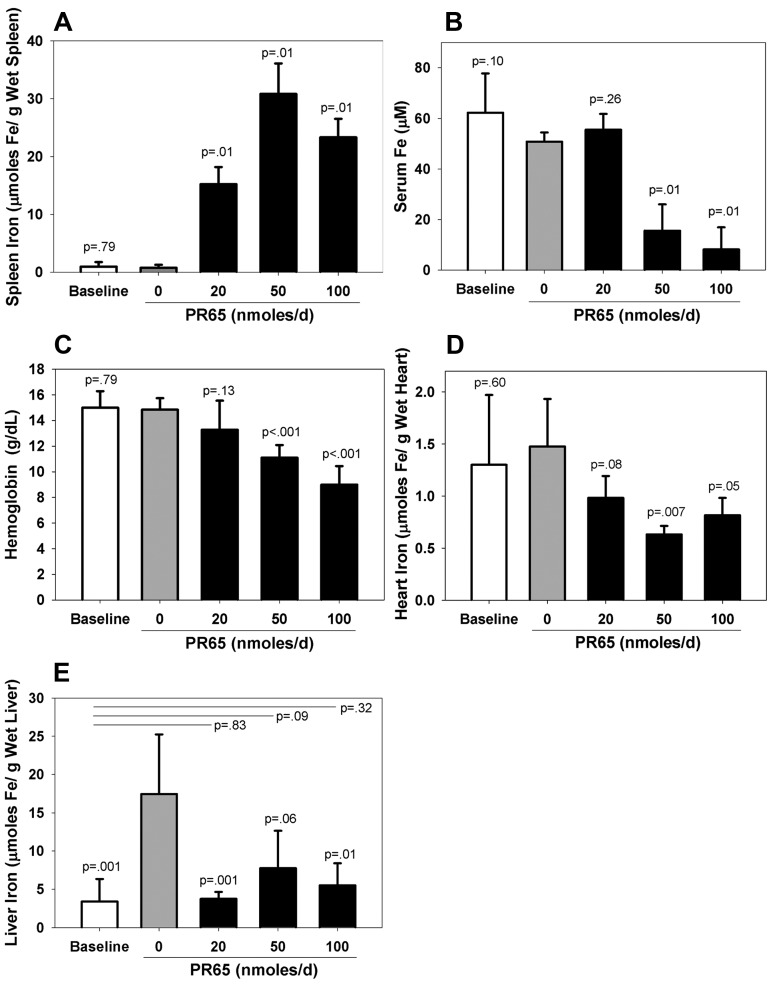
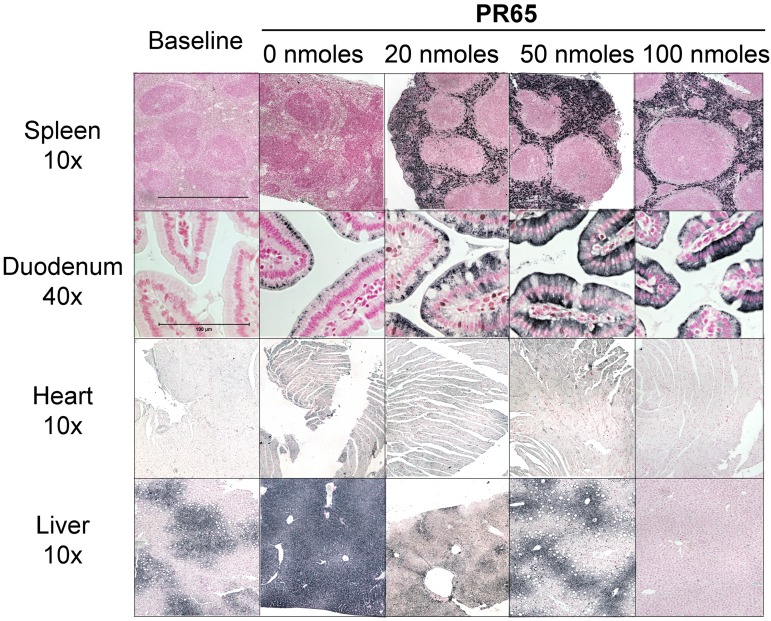
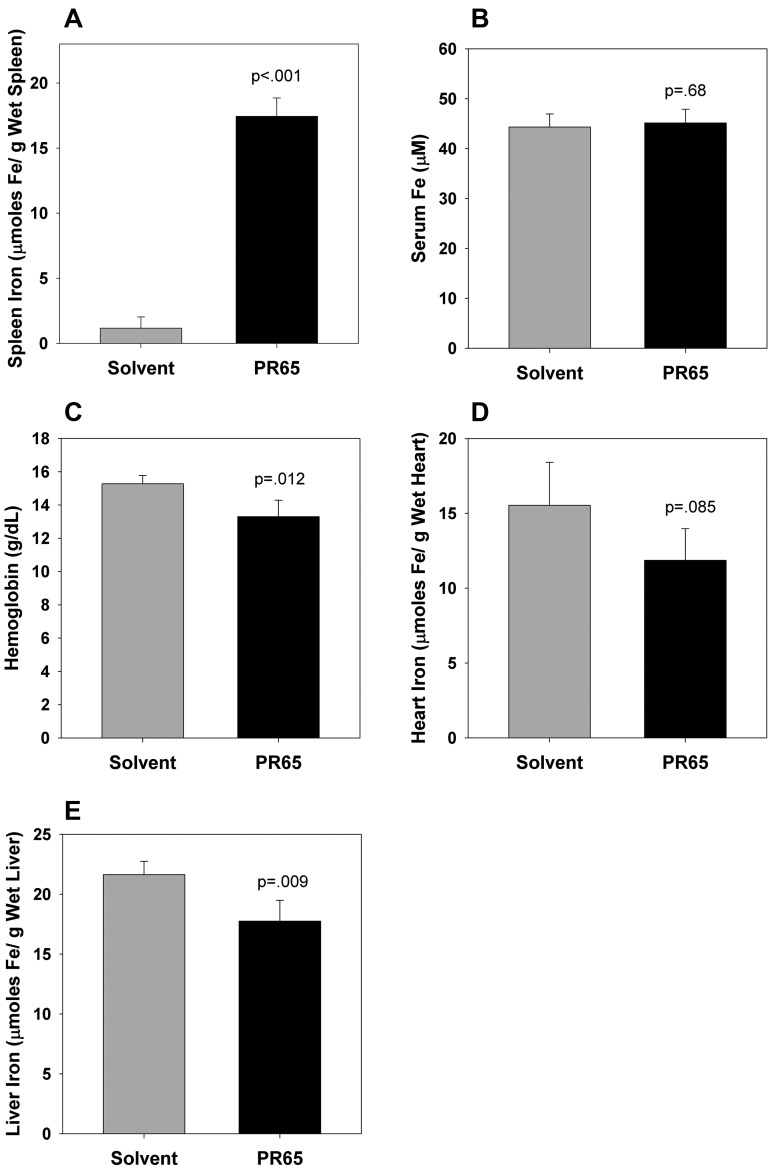
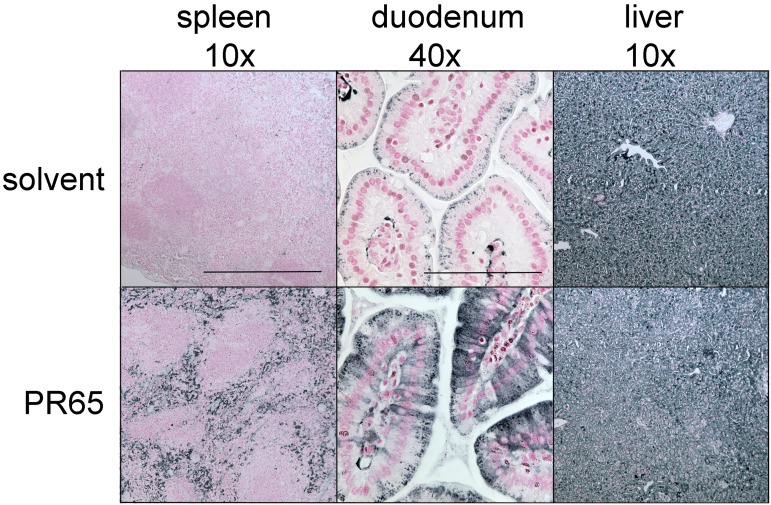
The deficiency of hepcidin, the hormone that controls iron absorption and its tissue distribution, is the cause of iron overload in nearly all forms of hereditary hemochromatosis and in untransfused iron-loading anemias. In a recent study, we reported the development of minihepcidins, small drug-like hepcidin agonists. Here we explore the feasibility of using minihepcidins for the prevention and treatment of iron overload in hepcidin-deficient mice. An optimized minihepcidin (PR65) was developed that had superior potency and duration of action compared with natural hepcidin or other minihepcidins, and favorable cost of synthesis. PR65 was administered by subcutaneous injection daily for 2 weeks to iron-depleted or iron-loaded hepcidin knockout mice. PR65 administration to iron-depleted mice prevented liver iron loading, decreased heart iron levels, and caused the expected iron retention in the spleen and duodenum. At high doses, PR65 treatment also caused anemia because of profound iron restriction. PR65 administration to hepcidin knockout mice with pre-existing iron overload had a more moderate effect and caused partial redistribution of iron from the liver to the spleen. Our study demonstrates that minihepcidins could be beneficial in iron overload disorders either used alone for prevention or possibly as adjunctive therapy with phlebotomy or chelation.
Figures
References
-
- Nemeth E, Tuttle MS, Powelson J, et al. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science. 2004;306(5704):2090–2093. - PubMed
-
- Brissot P, Bardou-Jacquet E, Jouanolle AM, Loreal O. Iron disorders of genetic origin: a changing world. Trends Mol Med. 2011;17(12):707–713. - PubMed
-
- Musallam KM, Cappellini MD, Wood JC, Taher AT. Iron overload in non-transfusion-dependent thalassemia: a clinical perspective. Blood Rev. 2012;26(Suppl 1):S16–S19. - PubMed
-
- Viatte L, Nicolas G, Lou DQ, et al. Chronic hepcidin induction causes hyposideremia and alters the pattern of cellular iron accumulation in hemochromatotic mice. Blood. 2006;107(7):2952–2958. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
