

Metallation and mismetallation of iron and manganese proteins in vitro and in vivo: the class I ribonucleotide reductases as a case study
- PMID: 22991063
- PMCID: PMC3488304
- DOI: 10.1039/c2mt20142a
Metallation and mismetallation of iron and manganese proteins in vitro and in vivo: the class I ribonucleotide reductases as a case study
Abstract
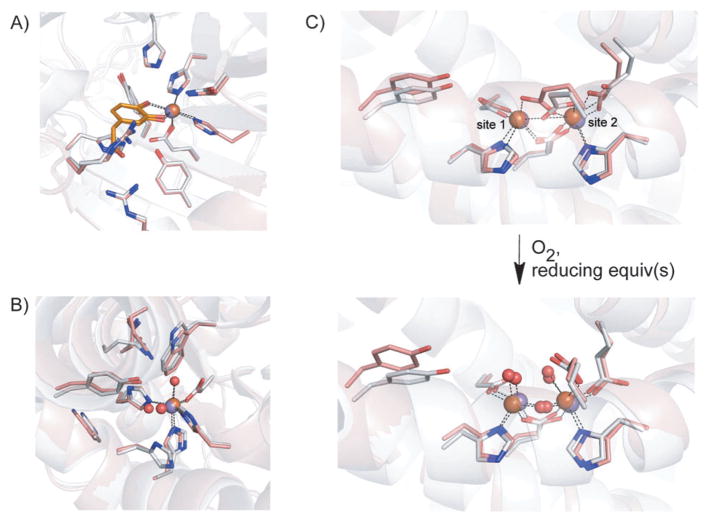
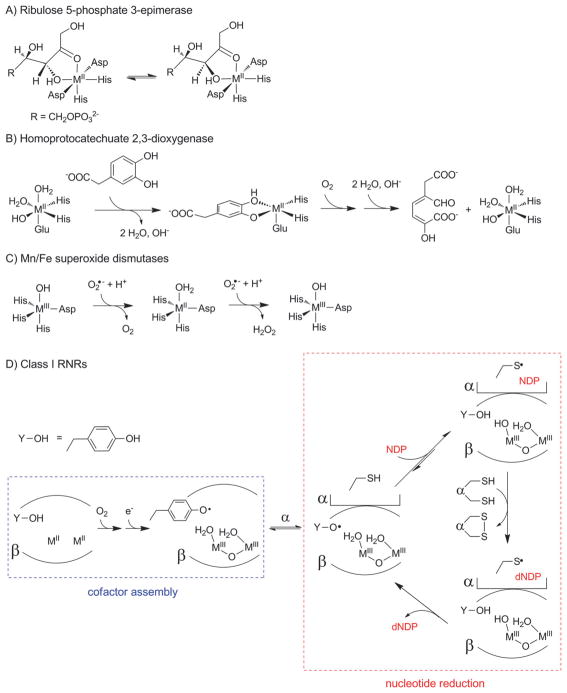
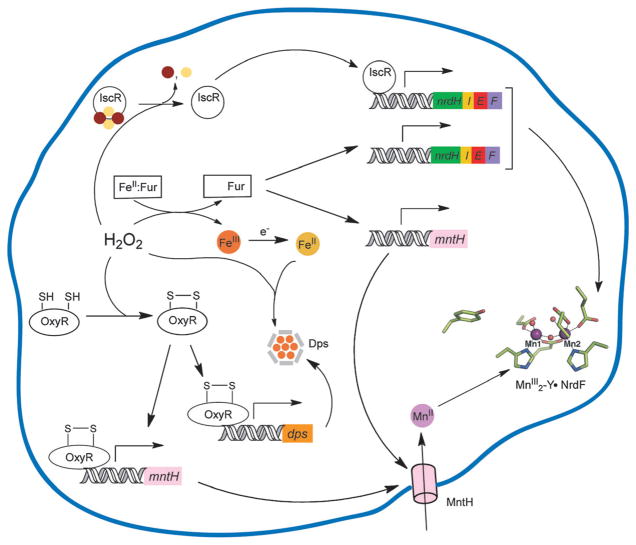
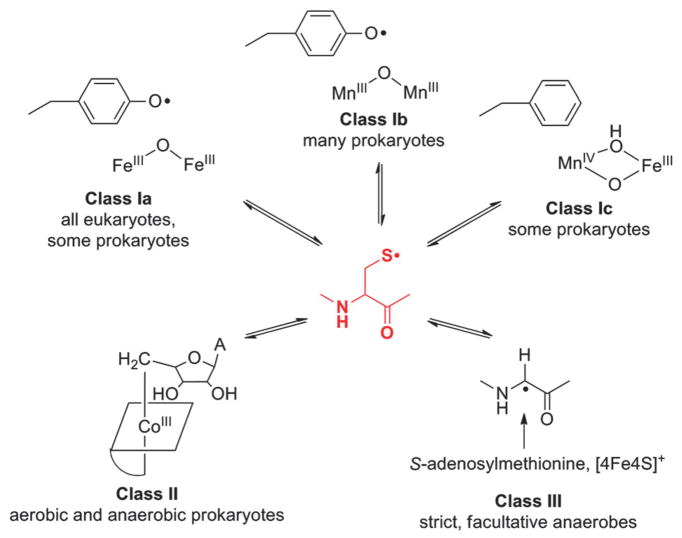
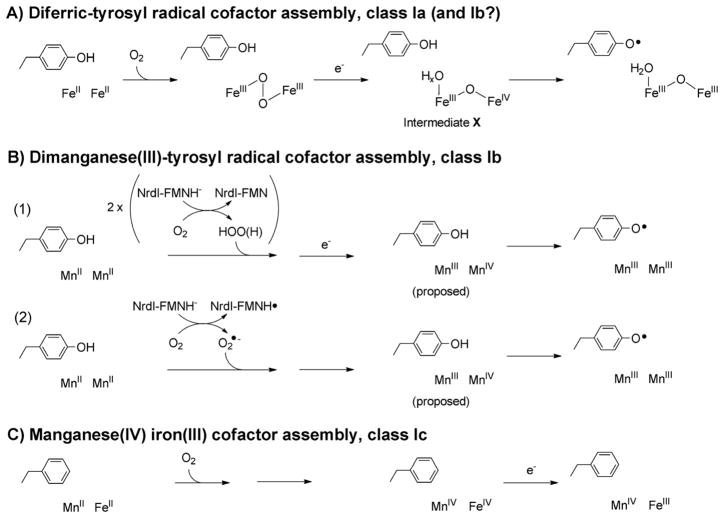
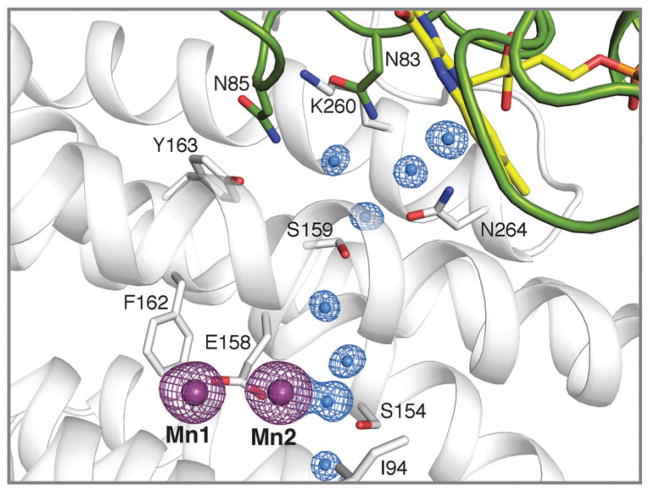
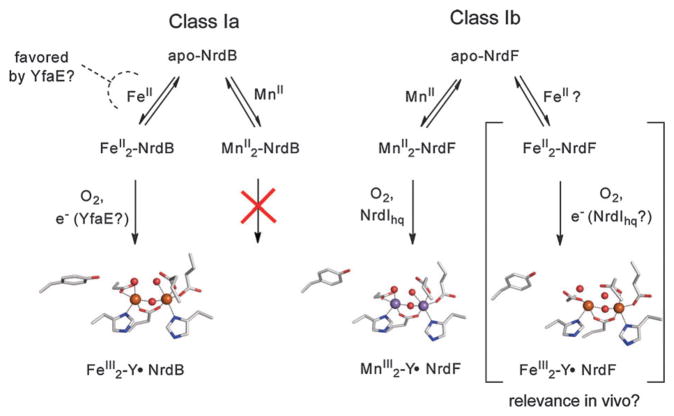
How cells ensure correct metallation of a given protein and whether a degree of promiscuity in metal binding has evolved are largely unanswered questions. In a classic case, iron- and manganese-dependent superoxide dismutases (SODs) catalyze the disproportionation of superoxide using highly similar protein scaffolds and nearly identical active sites. However, most of these enzymes are active with only one metal, although both metals can bind in vitro and in vivo. Iron(ii) and manganese(ii) bind weakly to most proteins and possess similar coordination preferences. Their distinct redox properties suggest that they are unlikely to be interchangeable in biological systems except when they function in Lewis acid catalytic roles, yet recent work suggests this is not always the case. This review summarizes the diversity of ways in which iron and manganese are substituted in similar or identical protein frameworks. As models, we discuss (1) enzymes, such as epimerases, thought to use Fe(II) as a Lewis acid under normal growth conditions but which switch to Mn(II) under oxidative stress; (2) extradiol dioxygenases, which have been found to use both Fe(II) and Mn(II), the redox role of which in catalysis remains to be elucidated; (3) SODs, which use redox chemistry and are generally metal-specific; and (4) the class I ribonucleotide reductases (RNRs), which have evolved unique biosynthetic pathways to control metallation. The primary focus is the class Ib RNRs, which can catalyze formation of a stable radical on a tyrosine residue in their β2 subunits using either a di-iron or a recently characterized dimanganese cofactor. The physiological roles of enzymes that can switch between iron and manganese cofactors are discussed, as are insights obtained from the studies of many groups regarding iron and manganese homeostasis and the divergent and convergent strategies organisms use for control of protein metallation. We propose that, in many of the systems discussed, "discrimination" between metals is not performed by the protein itself, but it is instead determined by the environment in which the protein is expressed.
Figures
References
-
- Mizuno K, Whittaker MM, Bachinger HP, Whittaker JW. J Biol Chem. 2004;279:27339–27344. - PubMed
-
- Tottey S, Waldron KJ, Firbank SJ, Reale B, Bessant C, Sato K, Cheek TR, Gray J, Banfield MJ, Dennison C, Robinson NJ. Nature. 2008;455:1138–1142. - PubMed
-
- Maret W. Metallomics. 2010;2:117–125. - PubMed
-
- Johnson DC, Dean DR, Smith AD, Johnston MK. Annu Rev Biochem. 2005;74:247–281. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
