

Review
doi: 10.1242/dev.080523.
Stomatal development: a plant's perspective on cell polarity, cell fate transitions and intercellular communication
Affiliations
- PMID: 22991435
- PMCID: PMC3445305
- DOI: 10.1242/dev.080523
Item in Clipboard
Review
Stomatal development: a plant's perspective on cell polarity, cell fate transitions and intercellular communication
Development.
2012 Oct.
Abstract
The plant stomatal lineage manifests features common to many developmental contexts: precursor cells are chosen from an initially equivalent field of cells, undergo asymmetric and self-renewing divisions, communicate among themselves and respond to information from a distance. As we review here, the experimental accessibility of these epidermal lineages, particularly in Arabidopsis, has made stomata a conceptual and technical framework for the study of cell fate, stem cells, and cell polarity in plants.
Figures
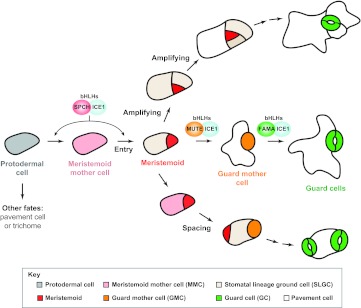
Cell fate transitions and divisions during Arabidopsis stomatal development. A protodermal cell (gray) commits to the stomatal lineage when it becomes a meristemoid mother cell (MMC; pink). MMCs undergo an asymmetric division (Entry) and produce a smaller meristemoid (red) and a larger stomatal lineage ground cell (SLGC; beige). Meristemoids may undergo additional self-renewing asymmetric divisions (Amplifying) or differentiate into a guard mother cell (GMC; orange). A GMC divides symmetrically once to form a pair of guard cells (GCs; green). SLGCs have two options: they can differentiate into pavement cells (white), or they can adopt an MMC fate and undergo an asymmetric division (Spacing) to create a new secondary meristemoid that is positioned away from the existing stomatal lineage cell. Three related bHLH transcription factors (bHLHs), SPCH, MUTE and FAMA, form heterodimers with bHLH-LZs ICE1 or SCRM2 (not shown), and are necessary and sufficient for driving the production of meristemoid, GMC and GC fates, respectively. For simplicity, they are shown at only one position in this figure, but they are also required during cell fate transitions in the amplifying and spacing branches.
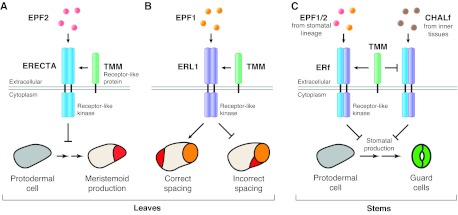
Ligand-receptor interactions regulate stomatal production and patterning. (A) In leaves, the secreted peptide EPF2 (pink) is produced by MMCs and early meristemoids. EPF2 is detected by the receptor-like kinase ERECTA (blue), present in protodermal cells (gray). In partnership with the receptor-like protein TMM (green), the EPF2-ERECTA pair is hypothesized to activate an intracellular signaling cascade that represses production of meristemoids (red). (B) EPF1 (orange, top), which is secreted by late meristemoids, GMCs or GCs, interacts with ERL1 (purple). The EPF1-ERL1 pair, together with TMM, induces signaling that affects the division plane such that the secondary meristemoid (red) forms away from pre-existing stoma or stomatal precursors. Illustrated here is a GMC (orange, bottom), and the results of correct and incorrect spacing of a newly formed secondary meristemoid. The EPF1-ERL1 pair also represses meristemoid differentiation (not shown). (C) In stems, ERf receptors (blue/purple) are subject to inadvertent activation by the EPF-related CHALf peptides (brown), which are normally produced in inner tissues for growth regulation. Stomatal lineage expression of TMM functions as a signaling insulator, repressing CHALf-mediated and promoting EPF1/2-mediated signaling. In A and B, ERECTA and ERL1 are shown as homodimers, but they may also form heterodimers with other members of the ERf.
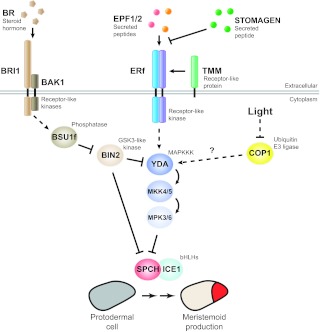
A mitogen-activated protein kinase (MAPK) pathway transduces and integrates intrinsic and environmental signals during stomatal production. A signal transaction cascade involving the MAPKKK YDA, MKK4/5 and MPK3/6 is employed to repress stomatal production. The EPFL-ERf-TMM module (including the antagonistic STOMAGEN) functioning within the stomatal lineage lies genetically upstream of YDA, activating it through an as yet unknown mechanism. The activated stomatal MAPK module can regulate stomatal development at multiple stages. Shown here is its repression of meristemoid production by MPK3/6 phosphorylation and downregulation of SPCH that is initiated upstream by the EPF2-ERECTA pair. The downstream targets of the MAPK pathway activated by the EPF1-ERL1 pair are not known. An intermediate signaling component in the brassinosteroid (BR) pathway, the GSK3-like kinase BIN2, interfaces with the stomatal development pathway at two levels: by phosphorylating and inhibiting YDA (thus increasing stomata) and by phosphorylating and repressing SPCH (thus decreasing stomata). Light also regulates the number of stomata through the ubiquitin E3 ligase COP1, a central repressor in light signal transduction. Genetic evidence has placed YDA downstream of COP1 but the nature of their molecular link is not known. Arrows indicate positive interactions, T-bars negative interactions. Solid lines indicate confirmed biochemical interactions and dashed lines are indirect or genetic interactions. BAK1, BRI1-ASSOCIATED RECEPTOR KINASE 1; BRI1, BR INSENSITIVE 1; BSU1f, family of BRI1-SUPPRESSOR 1.
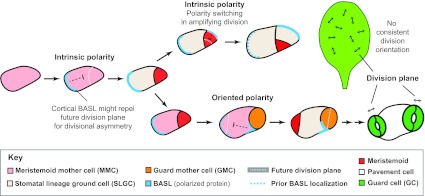
Asymmetric cell division and division polarity of stomata in Arabidopsis. The asymmetric divisions of MMCs with and without influences from stomatal lineage neighbors are referred to as having intrinsic and oriented polarity, respectively. Polarity is seen in cell sizes and division orientations, but also in the localization of the cortical proteins BASL (blue) and POLAR (not shown, but following the same trajectory as BASL). Prior to MMC asymmetric division, BASL localizes at the cortex of the MMC in crescents on the side at which the future SLGC will form. Models predict that such localization could generate cell polarity if the polarized protein acts to repel the nucleus and subsequent division plane. During amplifying divisions (top pathway), self-renewing meristemoids reorient cortical crescents away from the most recently formed walls, leading to a stomatal progenitor surrounded by lineally related non-stomatal sisters (see also Fig. 1, top pathway). White dotted lines indicate the future division plane and blue dotted lines indicate the previous position of BASL. In SLGCs adopting MMC fates (lower pathway), BASL must redistribute within the same cell into a crescent abutting the pre-existing meristemoid or GMC. For simplicity, the nuclear localization of BASL is not indicated. Although divisions of MMCs are precisely oriented relative to immediate neighbors, the divisions of MMCs and GMCs do not appear to be oriented relative to any landmarks or axes of the leaf.
References
-
- Abrash E. B., Bergmann D. C. (2010). Regional specification of stomatal production by the putative ligand CHALLAH. Development 137, 447-455 - PubMed
-
- Bergmann D. C., Sack F. D. (2007). Stomatal development. Annu. Rev. Plant Biol. 58, 163-181 - PubMed
-
- Bergmann D. C., Lukowitz W., Somerville C. R. (2004). Stomatal development and pattern controlled by a MAPKK kinase. Science 304, 1494-1497 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
