

Review
doi: 10.1038/nrm3440.
Epub 2012 Sep 20.
Endoplasmic reticulum-mitochondria contacts: function of the junction
Affiliations
- PMID: 22992592
- PMCID: PMC5111635
- DOI: 10.1038/nrm3440
Item in Clipboard
Review
Endoplasmic reticulum-mitochondria contacts: function of the junction
Nat Rev Mol Cell Biol.
2012 Oct.
Abstract
The most well-characterized organelle contact sites are those between the endoplasmic reticulum (ER) and mitochondria. Increased understanding is being gained of how ER-mitochondria contact sites are organized and which factors converge at this interface, some of which may provide a tethering function. The role of the ER-mitochondria junction in coordinating the functions of these two organelles is also becoming clearer, and it has been shown to be involved in the regulation of lipid synthesis, Ca(2+) signalling and the control of mitochondrial biogenesis and intracellular trafficking.
Figures
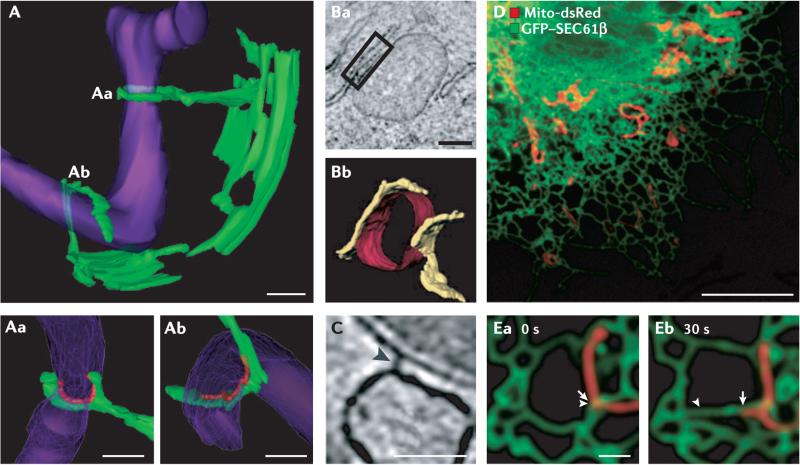
A | A three-dimensional (3D) electron microscope (EM) tomogram reveals contact sites (Aa and Ab) between the endoplasmic reticulum (ER; green) and a mitochondrion (purple) in a wild-type yeast cell. The mitochondrial membrane is constricted at position Aa where it is ‘clamped’ by an ER tubule. Higher-magnification images of contact sites Aa and Ab are shown in the bottom panels. Regions of contact are marked in red and are defined as points where the apposed membranes are within 30 nm of each other and free of ribosomes on the ER. B | An EM tomograph (Ba) and corresponding three-dimensional tomogram (Bb) of contact domains between the mitochondria (red) and the ER (yellow) in an inositol 1,4,5-trisphosphate receptor (IP3R) triple-knockout DT40 cultured chicken cell. The box in Ba shows a region of contact. C | An EM tomograph of a rat liver cell reveals electron-dense ‘tethers’ between the ER and mitochondrial membrane (marked by an arrowhead). D | A confocal fluorescent image of a Cos-7 cell labelled with mito-dsRed (showing the mitochondria in red) and green fluorescent protein (GFP)–SEC61β (showing the ER in green). E | A higher-magnification image of mitochondria and ER imaged live as in D at two time points. Arrows indicate the position of ER (arrowhead) and mitochondria (arrow) movement from 0 to 30 seconds. As the mitochondria moves, the ER moves with it. Scale bars represent 200 nm in A, 250 nm in B, 50 nm in C, 10 μM in D and 1 μM in E. Images in A are reproduced, with permission, from REF. © (2011) American Association for the Advancement of Science. Images in B and C are reproduced, with permission, from REF. © (2006) Rockefeller University Press. Images in D and E are reproduced, with permission, from REF. © (2010) Rockefeller University Press.
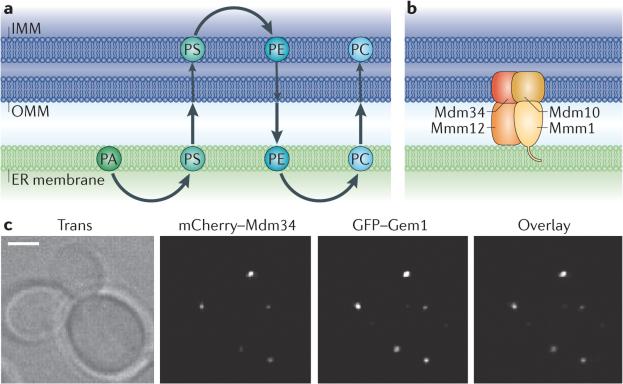
a | The biosynthesis pathway that generates phosphatidylcholine (PC) from phosphatidic acid (PA) requires sequential steps in both the endoplasmic reticulum (ER) and mitochondria. The enzymes that convert PA to phosphatidylserine (PS) or phosphatidylethanolamine (PE) to PC are in the ER, whereas the enzymes that convert PS to PE are in the mitochondria. So, to achieve the final lipid composition of either membrane, there must be a significant exchange of lipids between the two organelles. b | The ER–mitochondria encounter structure (ERMES) is a complex in yeast consisting of proteins that reside in both the ER and outer mitochondrial membranes (OMMs). ERMES forms a bridge between the ER and mitochondrial membranes: maintenance of mitochondrial morphology protein 1 (Mmm1) localizes to the ER; mitochondrial distribution and morphology protein 10 (Mdm10) and Mdm34 are in the OMM; and Mmm12 is cytoplasmic. c | The ERMES component Mdm34 (labelled with mCherry) localizes to punctate structures on mitochondria that colocalize with Gem1 (GTPase EF-hand protein of mitochondria 1). GFP, green fluorescent protein; IMM, inner mitochondrial membrane. Images in c are reproduced, with permission, from REF © (2011) National Academy of Sciences.
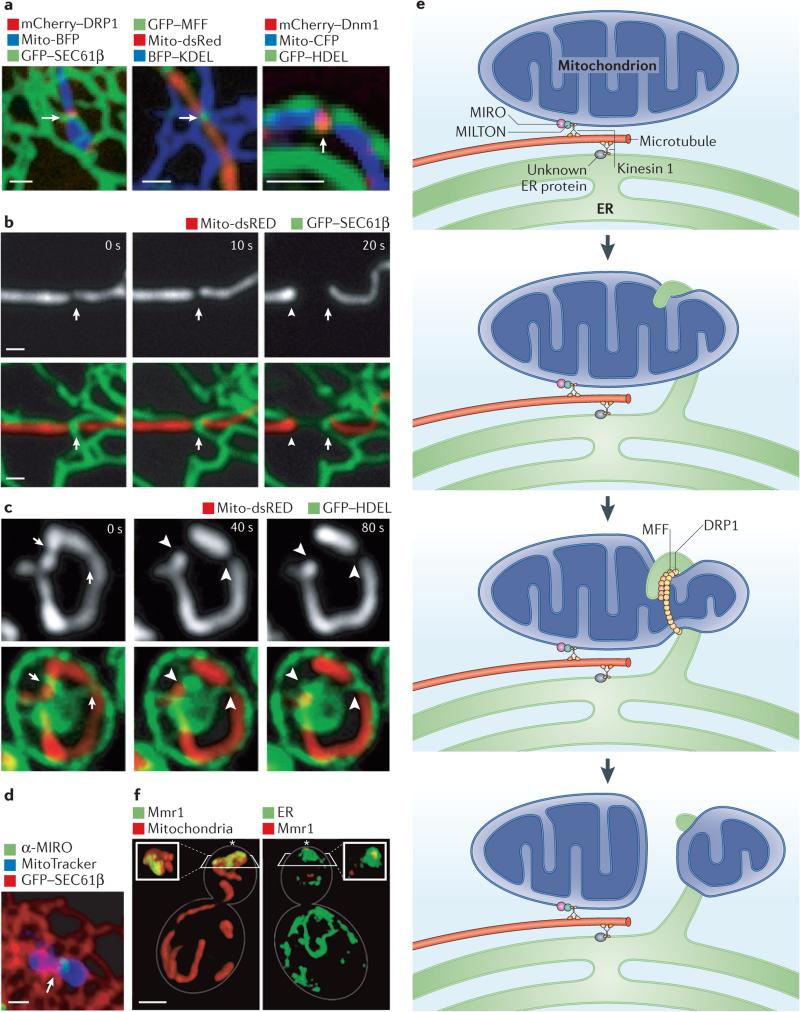
a | Confocal microscopy images showing the mitochondrial division machinery proteins DRP1 (dynamin-related protein 1) and MFF (mitochondrial fission factor) in Cos-7 cells and the yeast division machinery dynamin Dnm1 localizing to positions where endoplasmic reticulum (ER) tubules circumscribe the mitochondrial membrane. b,c | Live confocal fluorescence time-lapse images of Cos-7 cells (b) and yeast cells (c) showing mitochondrial constriction followed by division taking place at the site of an ER tubule crossing the mitochondria. Arrows indicate the initial site of constriction and arrowheads indicate the site of mitochondrial division, d | The Ca2+ binding protein MIRO (mitochondrial Rho GTPase), which regulates mitochondrial motility, localizes to a position (indicated by an arrow) where an ER tubule crosses over the mitochondrial membrane in a Cos-7 cell, e | A model of multiple factors that converge at ER–mitochondria contact sites to regulate mitochondrial dynamics. Both mitochondria and the ER are linked to microtubules by factors that associate with the microtubule motor kinesin 1. In the case of mitochondria, this occurs through kinesin 1 binding to the cytoplasmic protein MILTON, which in turn binds MIRO on the outer mitochondrial membrane. MIRO is important for mitochondrial movement. Less is known about the ER proteins that tether the ER to microtubules. Mitochondrial constriction mediated by DRP1 occurs at sites of ER–mitochondria contact, and one possibility is that ER contact promotes initial constriction of the mitochondria before DRP1 recruitment. Ultimately, mitochondrial fission is promoted by DRP1 and its cofactor MFF. f | Volume rendering of structured illumination microscopy images show a yeast cell during coordinated organelle inheritance into the bud. This is mediated by contact sites between the ER and the mitochondria, and normal ER inheritance is important for mitochondrial inheritance. Mitochondrial MYO2 receptor-related protein 1 (Mmr1) links the mitochondria to the ER during inheritance and accumulates between the mitochondria (shown in red) and the cortical ER (labelled by green fluorescent protein (GFP)–Sec63, in green). The bud tip is indicated by an asterisk. Scale bars represent 1 μM. Images in a–c are reproduced, with permission, from REF. © (2011) American Association for the Advancement of Science. The image in d is reproduced, with permission, from REF. © (2011) National Academy of Sciences. Images in f are reproduced, with permission, from REF. © (2011) Elsevier. BFP, blue fluorescent protein; CFP, cyan fluorescent protein; HDEL, His-Asp-Glu-Leu; KDEL, Lys-Asp-Glu-Leu.
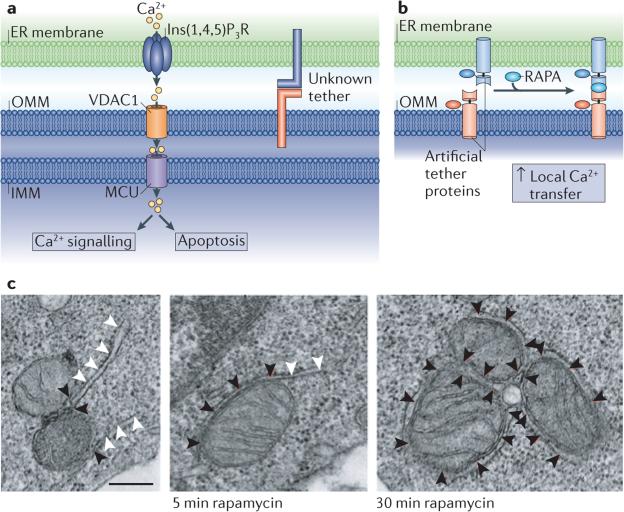
a | Ca2+ transfer is proposed to occur from the endoplasmic reticulum (ER) lumen into the mitochondria at contact sites. This requires the inositol 1,4,5-trisphosphate receptor(Ins(1,4,5)P3R) on the ER membrane, and Ca2+ uptake is thought to be mediated by voltage-dependent anion selective channel protein 1 (VDAC1) on the outer mitochondrial membrane (OMM). More recently, the mitochondrial calcium uniporter (MCU) has been identified as the regulator of Ca2+ uptake at the inner mitochondrial membrane (IMM), and this is likely to require Ca2+ concentrations found near ER–mitochondria contacts. b | A rapamycin-inducible tether has demonstrated the importance of tethering between the ER and mitochondria for Ca2+ transfer. Half of the tether is localized to the OMM by fusing a mitochondrial localization signal (taken from mitochondrial A-kinase anchor protein 1 (AKAP1), residues 34–63) to 12 kDa FK506-binding protein (FKBP12)–mitochondrial red fluorescent protein 1 (mRFP1) (red). The partner protein is targeted to the ER membrane using an ER targeting signal (taken from SAC1, residues 521–587) fused to FKBP12 rapamycin binding domain (FRB)–cyan fluorescent protein (CFP) (blue). Treatment with rapamycin (RAPA) induces dimerization between FKBP12 and FRB, and thus membrane tethering; this increases local Ca2+ transfer. c | An electron microscope tomograph of RBL-2H3 cells expressing the artificial imaged tether before and after rapamycin-induced dimerization. Black arrowheads indicate ER–mitochondria contact, and white arrowheads indicate ER not in contact with mitochondria. Scale bar represents 250 nm. Images in b and c are reproduced, with permission, from REF. © (2010) Elsevier.
References
-
- Elbaz Y, Schuldiner M. Staying in touch: the molecular era of organelle contact sites. Trends Biochem. Sci. 2011;36:616–623. - PubMed
-
- Friedman JR, et al. ER tubules mark sites of mitochondrial division. Science. 2011;334:358–362. [Demonstrates that mitochondrial division occurs at positions where ER tubules contact mitochondria, and that contact and constriction occurs prior to recruitment of the division machinery.] - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
