

Direct tests of enzymatic heme degradation by the malaria parasite Plasmodium falciparum
- PMID: 22992734
- PMCID: PMC3488054
- DOI: 10.1074/jbc.M112.414078
Direct tests of enzymatic heme degradation by the malaria parasite Plasmodium falciparum
Abstract
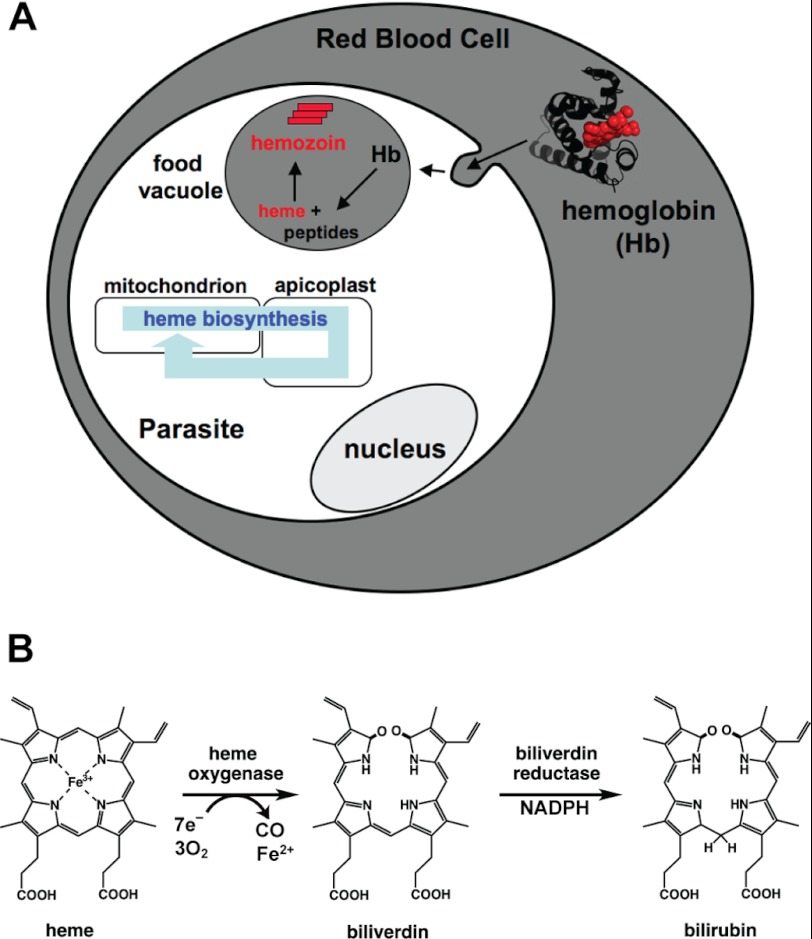
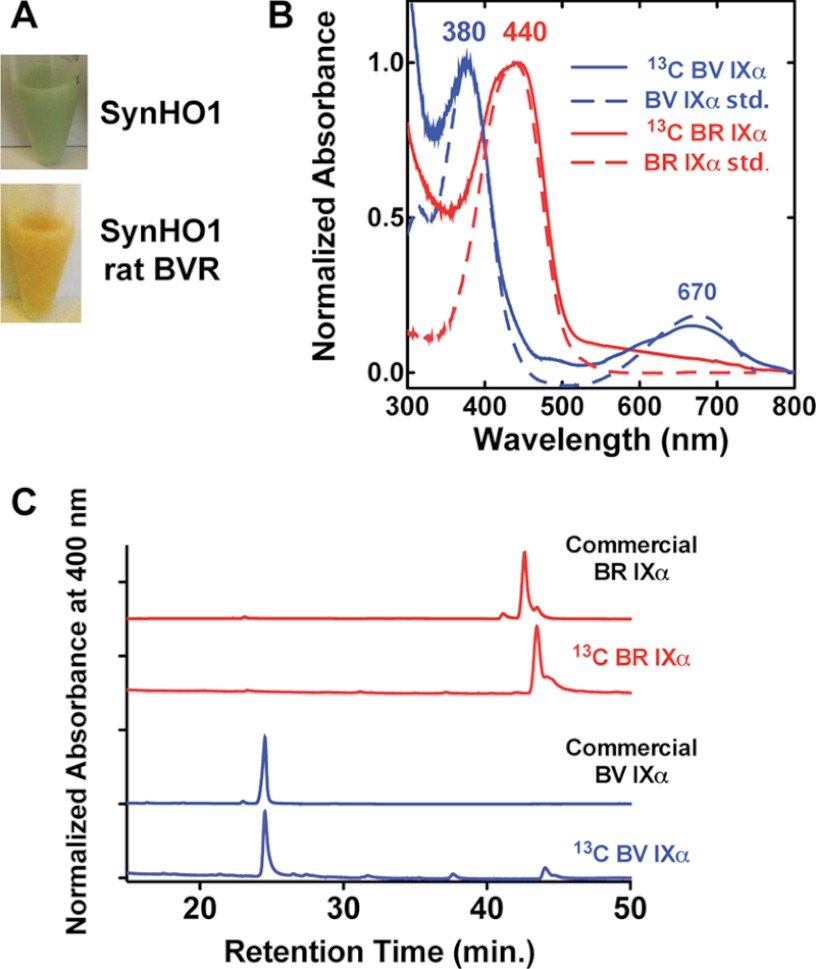
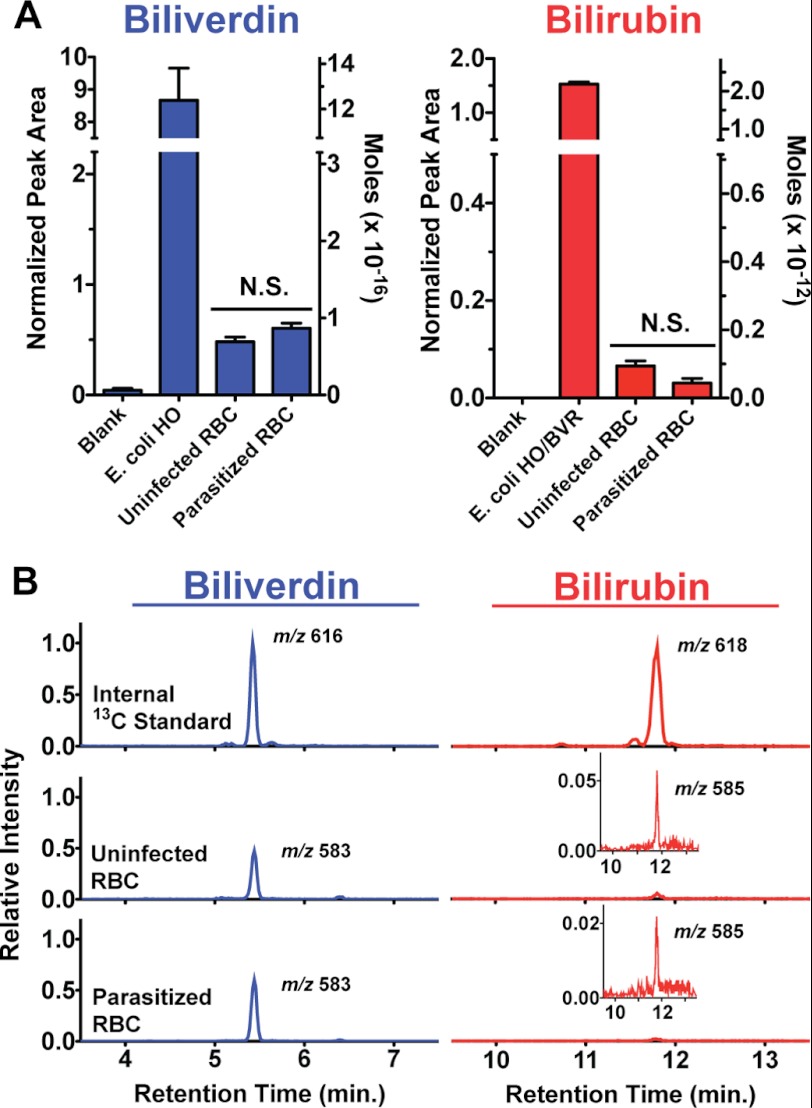
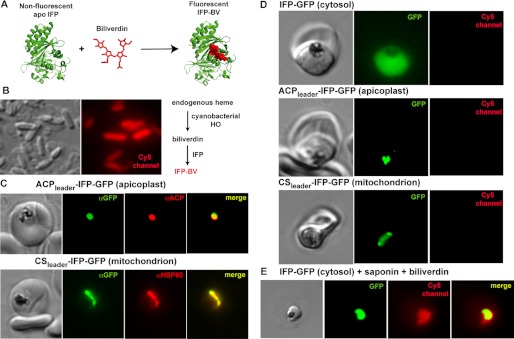
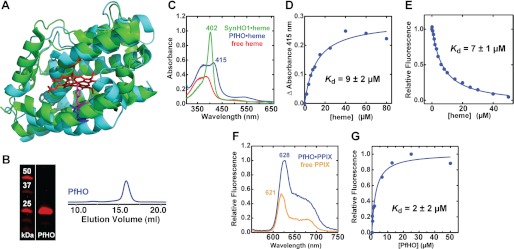
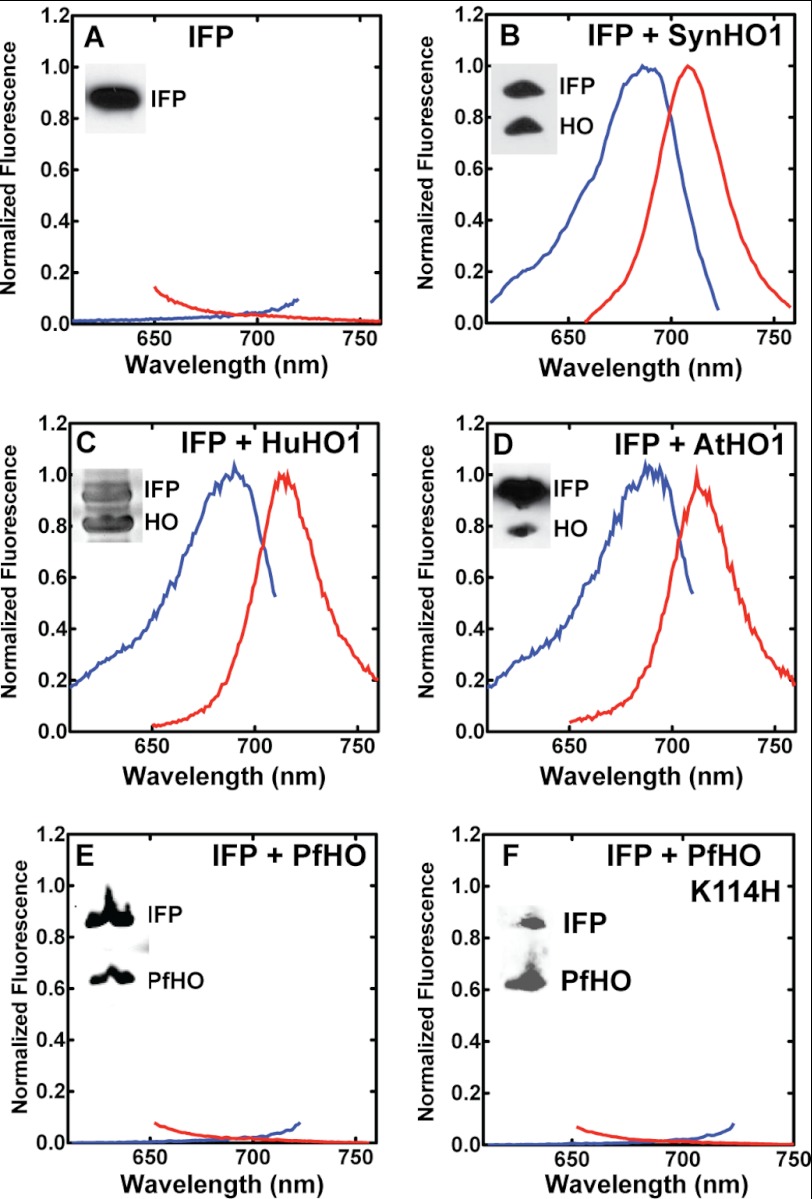
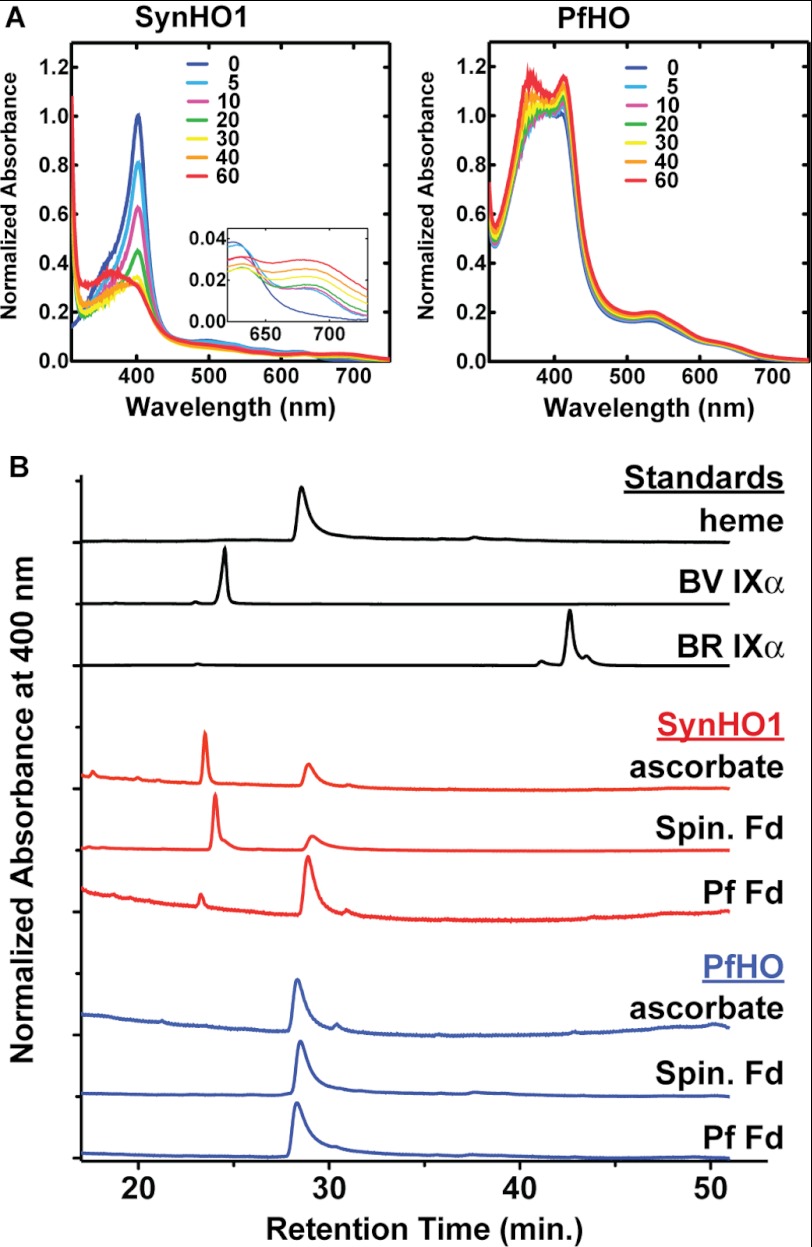
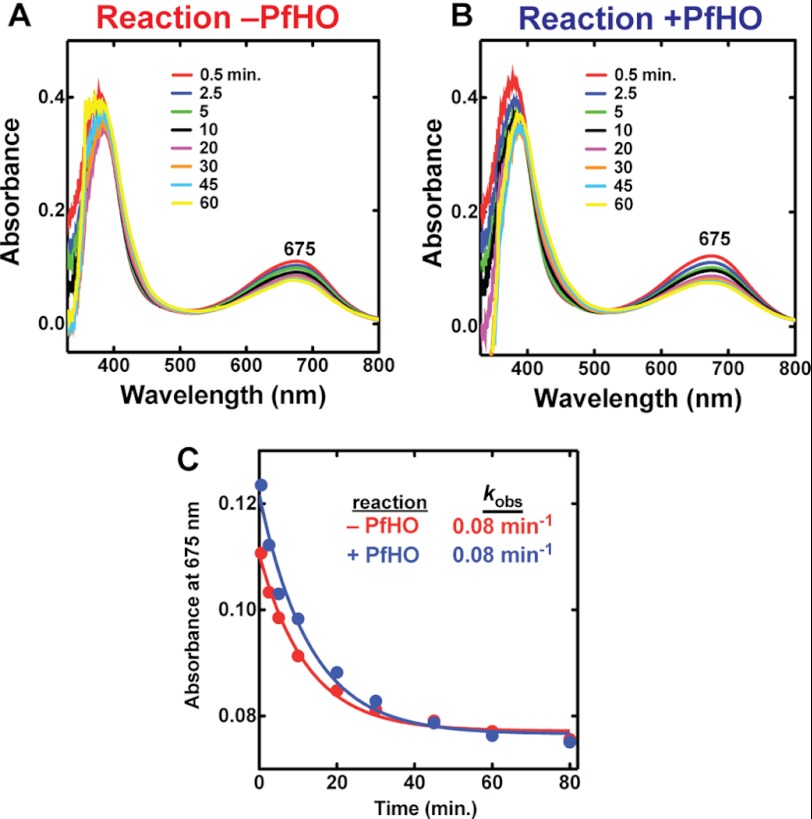
Malaria parasites generate vast quantities of heme during blood stage infection via hemoglobin digestion and limited de novo biosynthesis, but it remains unclear if parasites metabolize heme for utilization or disposal. Recent in vitro experiments with a heme oxygenase (HO)-like protein from Plasmodium falciparum suggested that parasites may enzymatically degrade some heme to the canonical HO product, biliverdin (BV), or its downstream metabolite, bilirubin (BR). To directly test for BV and BR production by P. falciparum parasites, we DMSO-extracted equal numbers of infected and uninfected erythrocytes and developed a sensitive LC-MS/MS assay to quantify these tetrapyrroles. We found comparable low levels of BV and BR in both samples, suggesting the absence of HO activity in parasites. We further tested live parasites by targeted expression of a fluorescent BV-binding protein within the parasite cytosol, mitochondrion, and plant-like plastid. This probe could detect exogenously added BV but gave no signal indicative of endogenous BV production within parasites. Finally, we recombinantly expressed and tested the proposed heme degrading activity of the HO-like protein, PfHO. Although PfHO bound heme and protoporphyrin IX with modest affinity, it did not catalyze heme degradation in vivo within bacteria or in vitro in UV absorbance and HPLC assays. These observations are consistent with PfHO's lack of a heme-coordinating His residue and suggest an alternative function within parasites. We conclude that P. falciparum parasites lack a canonical HO pathway for heme degradation and thus rely fully on alternative mechanisms for heme detoxification and iron acquisition during blood stage infection.
Figures

References
-
- Francis S. E., Sullivan D. J., Jr., Goldberg D. E. (1997) Hemoglobin metabolism in the malaria parasite Plasmodium falciparum. Annu. Rev. Microbiol. 51, 97–123 - PubMed
-
- van Dooren G. G., Kennedy A. T., McFadden G. I. (2012) The use and abuse of heme in apicomplexan parasites. Antioxid. Redox Signal. 17, 634–656 - PubMed
-
- Surolia N., Padmanaban G. (1992) De novo biosynthesis of heme offers a new chemotherapeutic target in the human malarial parasite. Biochem. Biophys. Res. Commun. 187, 744–750 - PubMed
-
- Nagaraj V. A., Prasad D., Rangarajan P. N., Padmanaban G. (2009) Mitochondrial localization of functional ferrochelatase from Plasmodium falciparum. Mol. Biochem. Parasitol. 168, 109–112 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HD001459/HD/NICHD NIH HHS/United States
- HL101263/HL/NHLBI NIH HHS/United States
- P60-DK20579/DK/NIDDK NIH HHS/United States
- P50 DK064540/DK/NIDDK NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- P30 HL101263/HL/NHLBI NIH HHS/United States
- DK064540/DK/NIDDK NIH HHS/United States
- K12 HD001459/HD/NICHD NIH HHS/United States
- P60 DK020579/DK/NIDDK NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- P41 RR000954/RR/NCRR NIH HHS/United States
- P41-RR00954/RR/NCRR NIH HHS/United States
- P30-DK56341/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
