

Tbx18 targets dermal condensates for labeling, isolation, and gene ablation during embryonic hair follicle formation
- PMID: 22992803
- PMCID: PMC3530628
- DOI: 10.1038/jid.2012.329
Tbx18 targets dermal condensates for labeling, isolation, and gene ablation during embryonic hair follicle formation
Abstract
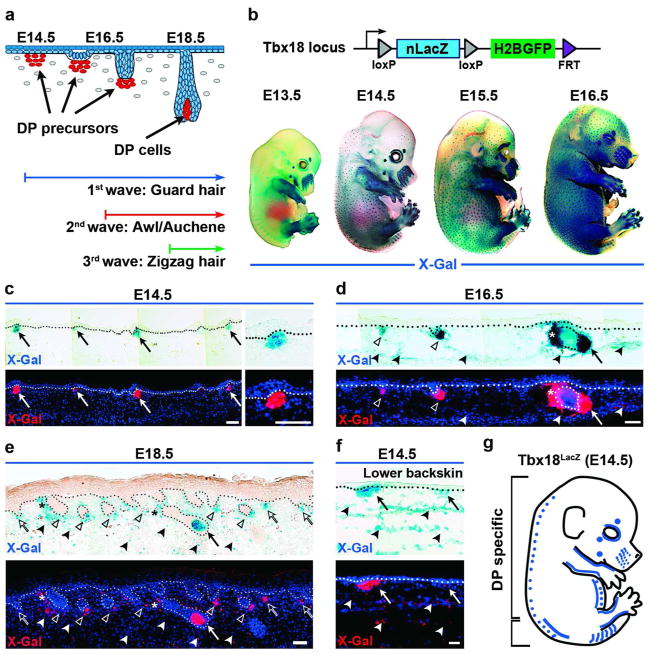
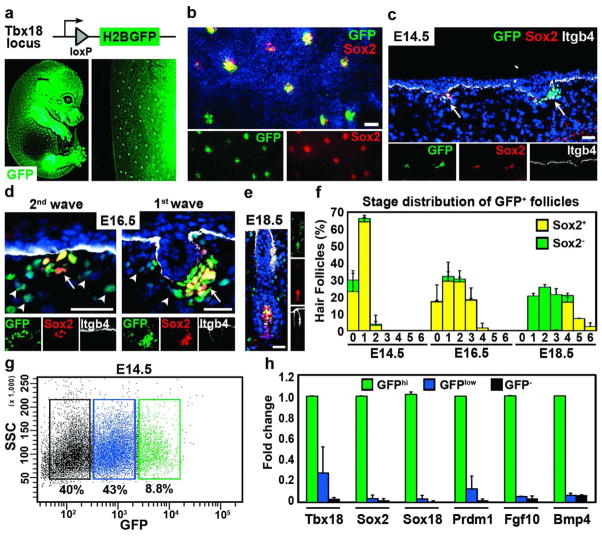
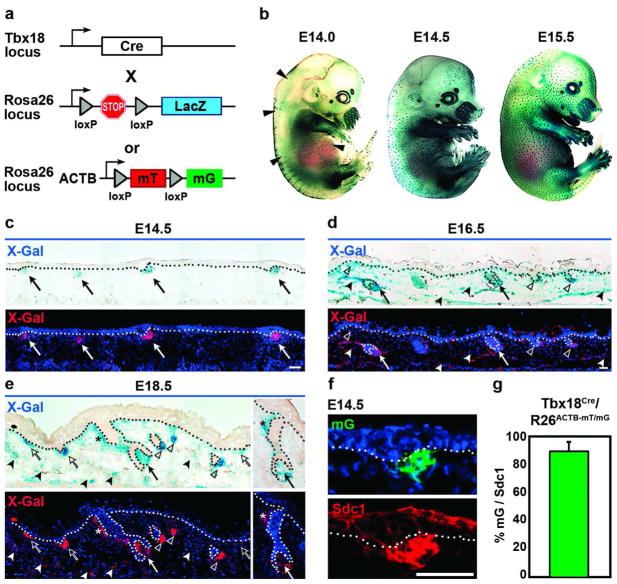
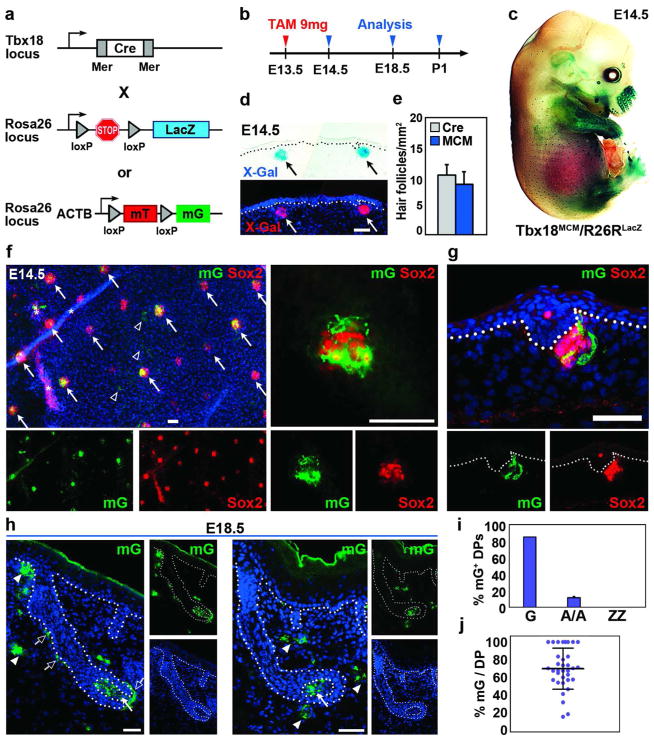
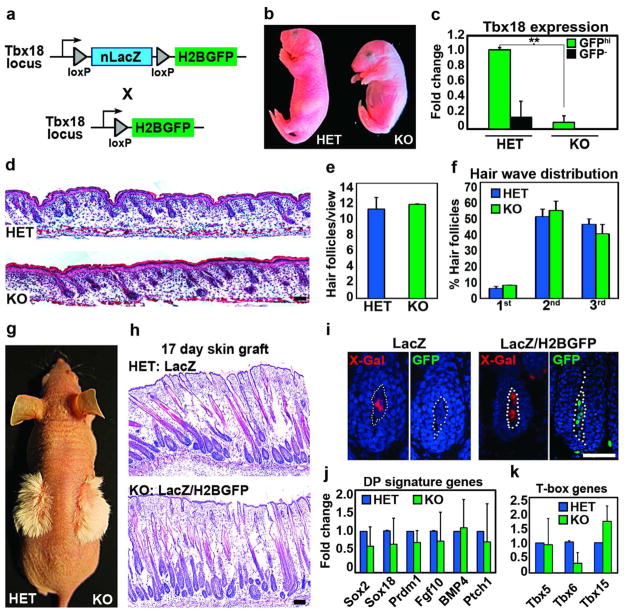
How cell fate decisions of stem and progenitor cells are regulated by their microenvironment or niche is a central question in stem cell and regenerative biology. Although functional analysis of hair follicle epithelial stem cells by gene targeting is well established, the molecular and genetic characterization of the dermal counterpart during embryonic morphogenesis has been lacking because of the absence of cell type-specific drivers. Here, we report that T-box transcription factor Tbx18 specifically marks dermal papilla (DP) precursor cells during embryonic hair follicle morphogenesis. With Tbx18(LacZ), Tbx18(H2BGFP), and Tbx18(Cre) knock-in mouse models, we demonstrate LacZ and H2BGFP (nuclear green fluorescent protein) expression and Cre activity in dermal condensates of nascent first-wave hair follicles at E14.5. As Tbx18 expression becomes more widespread throughout the dermis at later developmental stages, we use tamoxifen-inducible Cre-expressing mice, Tbx18(MerCreMer), to exclusively target DP precursor cells and their progeny. Finally, we ablate Tbx18 in full knockout mice, but find no perturbations in hair follicle formation, suggesting that Tbx18 is dispensable for normal DP function. In summary, our study establishes Tbx18 as a genetic driver to target for the first time embryonic DP precursors for labeling, isolation, and gene ablation that will greatly enhance investigations into their molecular functions during hair follicle morphogenesis.
Conflict of interest statement
The authors state no conflict of interest.
Figures
References
-
- Andl T, Reddy ST, Gaddapara T, Millar SE. WNT signals are required for the initiation of hair follicle development. Dev Cell. 2002;2:643–53. - PubMed
-
- Botchkarev VA, Botchkareva NV, Roth W, Nakamura M, Chen LH, Herzog W, et al. Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat Cell Biol. 1999;1:158–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
