

Gating reaction mechanism of neuronal NMDA receptors
- PMID: 22993263
- PMCID: PMC3544869
- DOI: 10.1152/jn.00551.2012
Gating reaction mechanism of neuronal NMDA receptors
Abstract
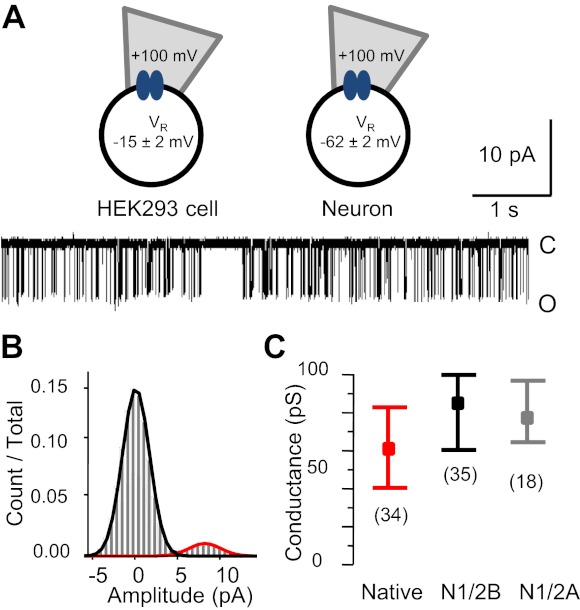
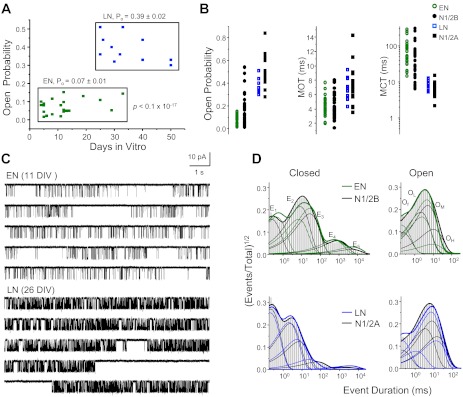
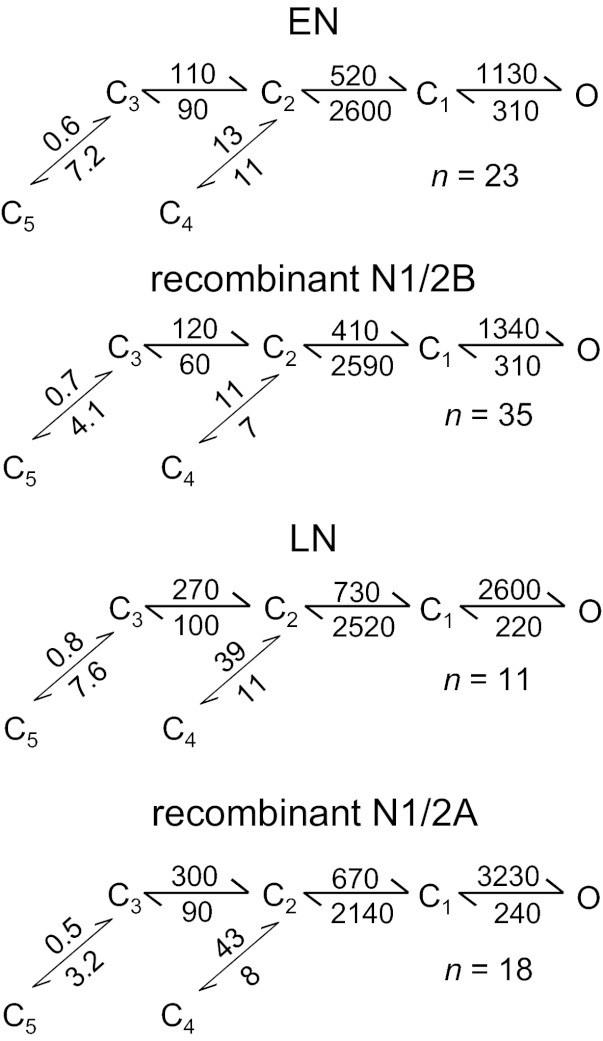
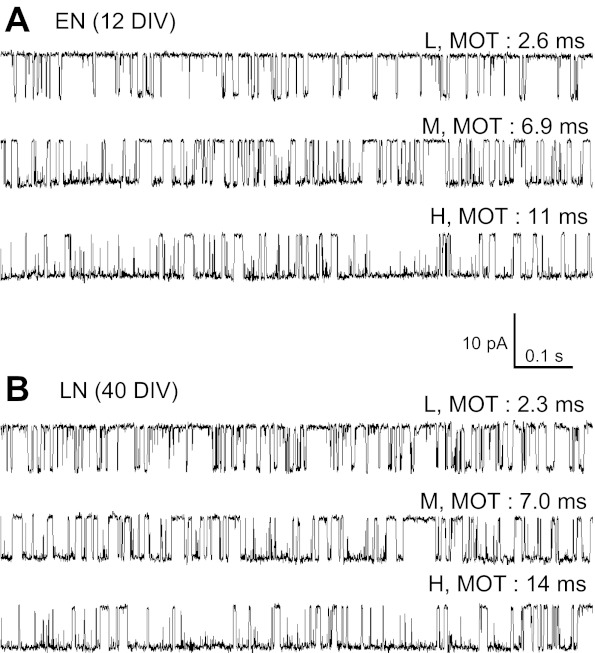
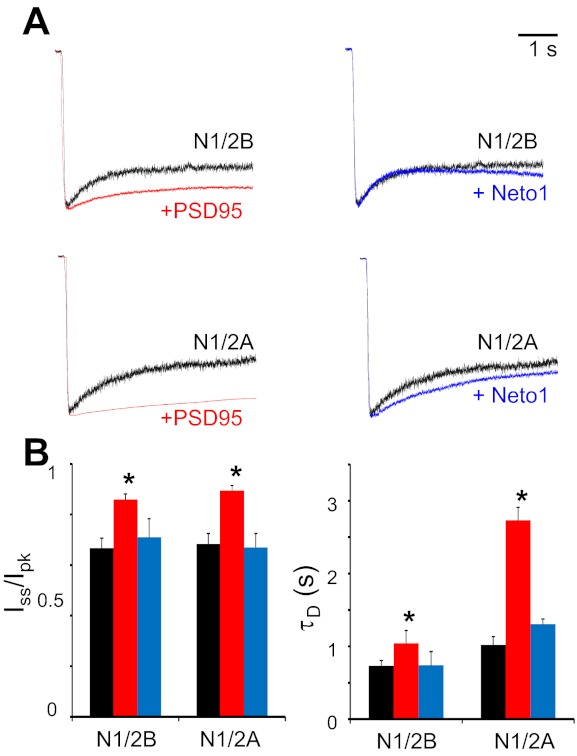
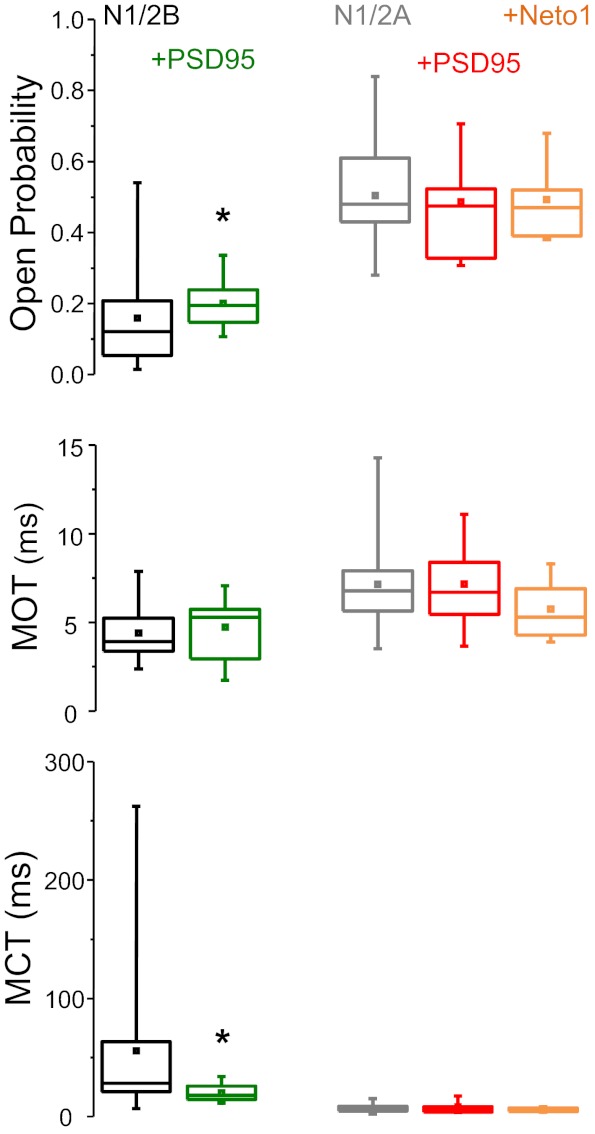
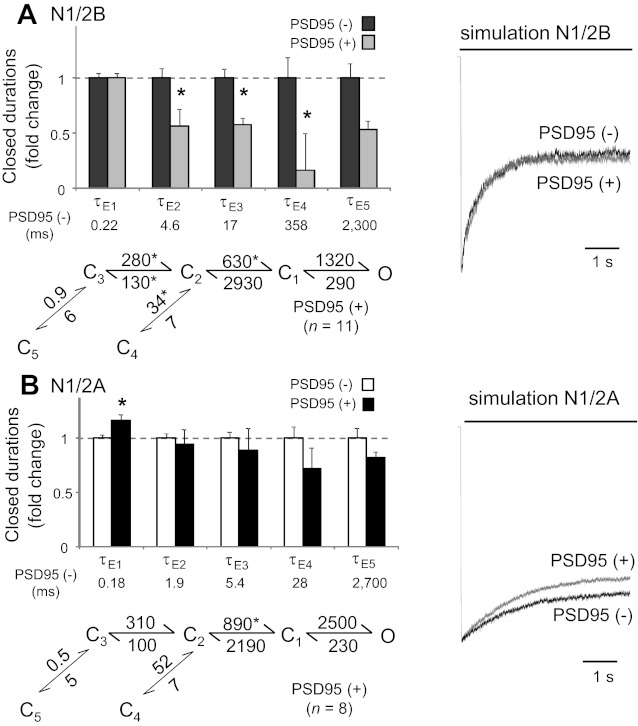
The activation mechanisms of recombinant N-methyl-d-aspartate receptors (NRs) have been established in sufficient detail to account for their single channel and macroscopic responses; however, the reaction mechanism of native NRs remains uncertain due to indetermination of the isoforms expressed and possible neuron-specific factors. To delineate the activation mechanism of native NRs, we examined the kinetic properties of currents generated by individual channels located at the soma of cultured rat neurons. Cells were dissociated from the embryonic cerebral cortex or hippocampus, and on-cell single channel recordings were done between 4 and 50 days in vitro (DIV). We observed two types of kinetics that correlated with the age of the culture. When we segregated recordings by culture age, we found that receptors recorded from early (4-33 DIV) and late (25-50 DIV) cultures had smaller unitary conductances but had kinetic profiles that matched closely those of recombinant 2B- or 2A-containing receptors, respectively. In addition, we examined the effects of cotransfection with postsynaptic density protein 95 or neuropilin tolloid-like protein 1 on recombinant receptors expressed in human embryonic kidney-293 cells. Our results add support to the view that neuronal cultures recapitulate the developmental patterns of receptor expression observed in the intact animal and demonstrate that the activation mechanism of somatic neuronal NRs is similar to that described for recombinant receptors of defined subunit composition.
Figures
References
-
- Banke TG, Traynelis SF. Activation of NR1/NR2B NMDA receptors. Nat Neurosci 6: 144–152, 2003 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials