

The human brain intracerebral microvascular system: development and structure
- PMID: 22993505
- PMCID: PMC3440694
- DOI: 10.3389/fnana.2012.00038
The human brain intracerebral microvascular system: development and structure
Abstract
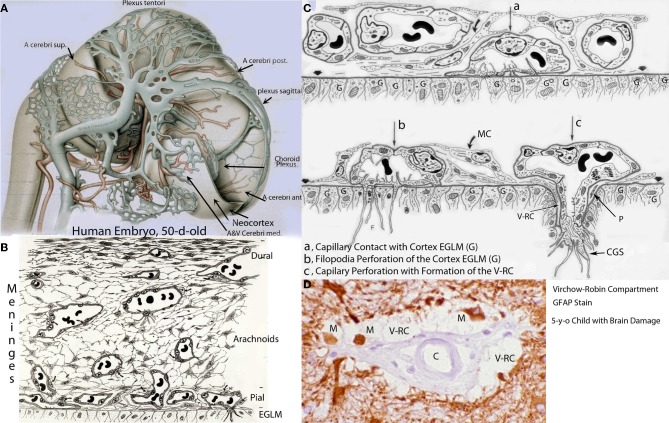
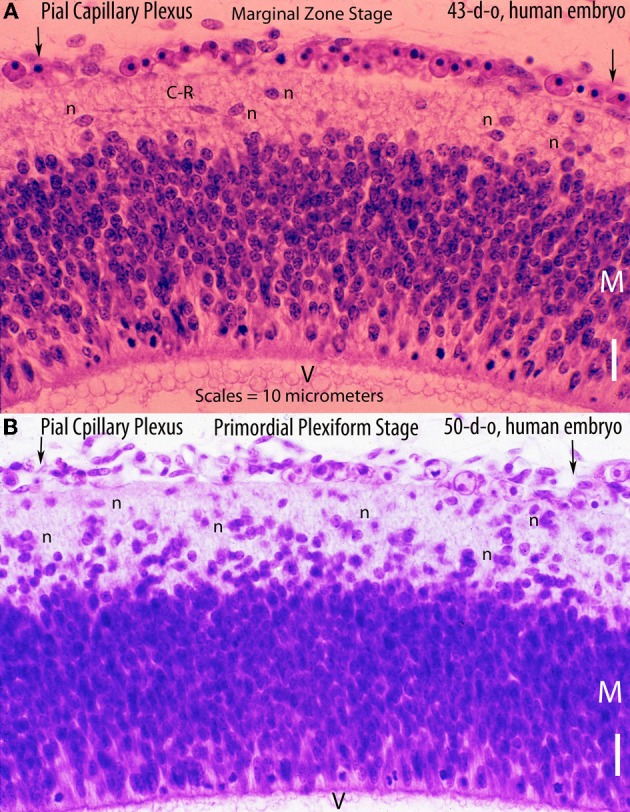
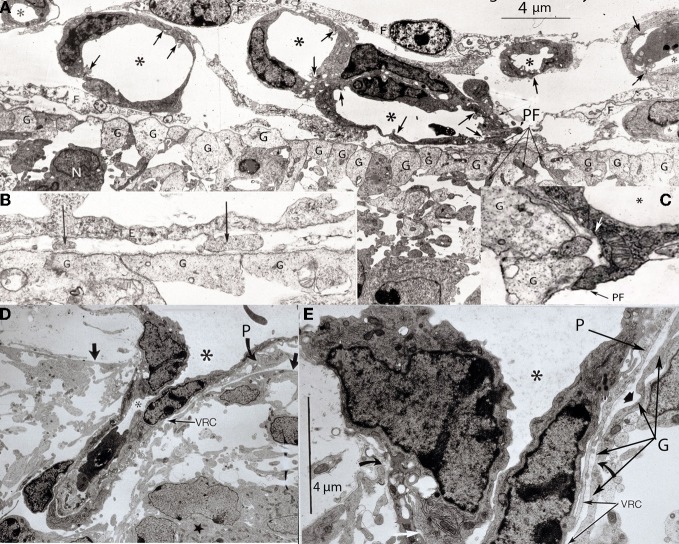
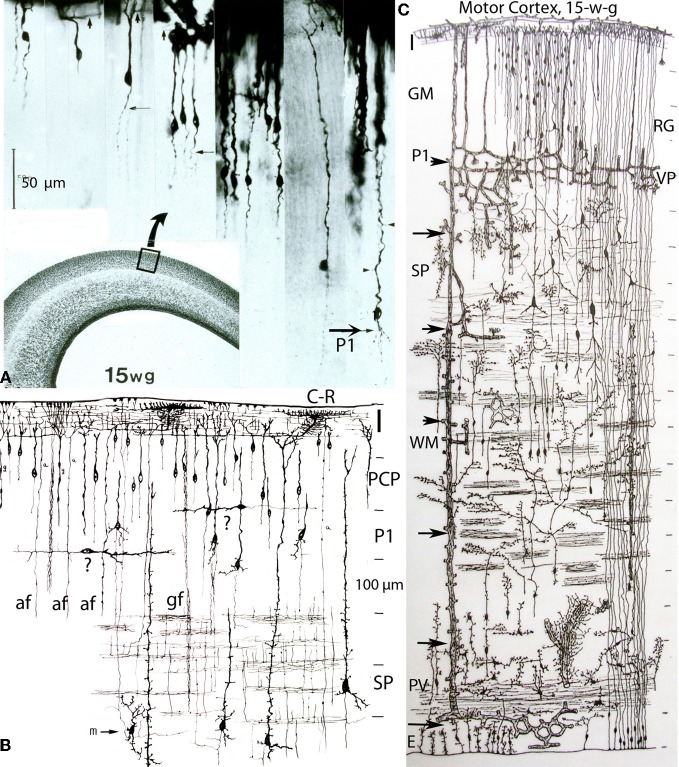
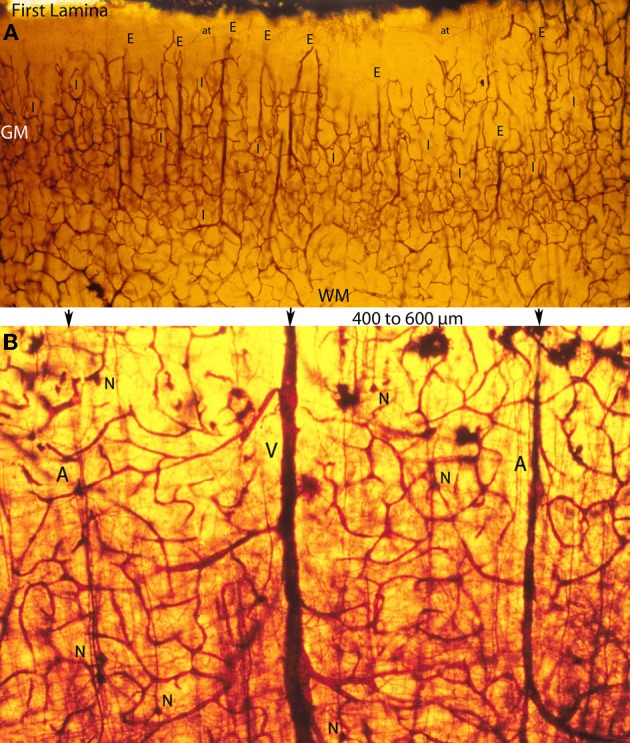
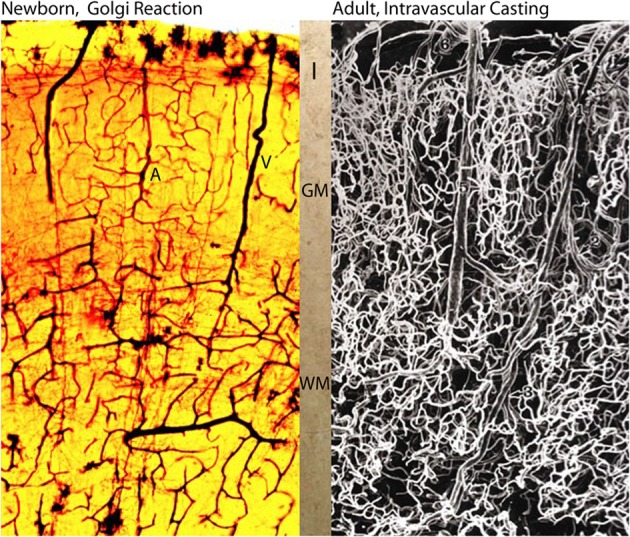
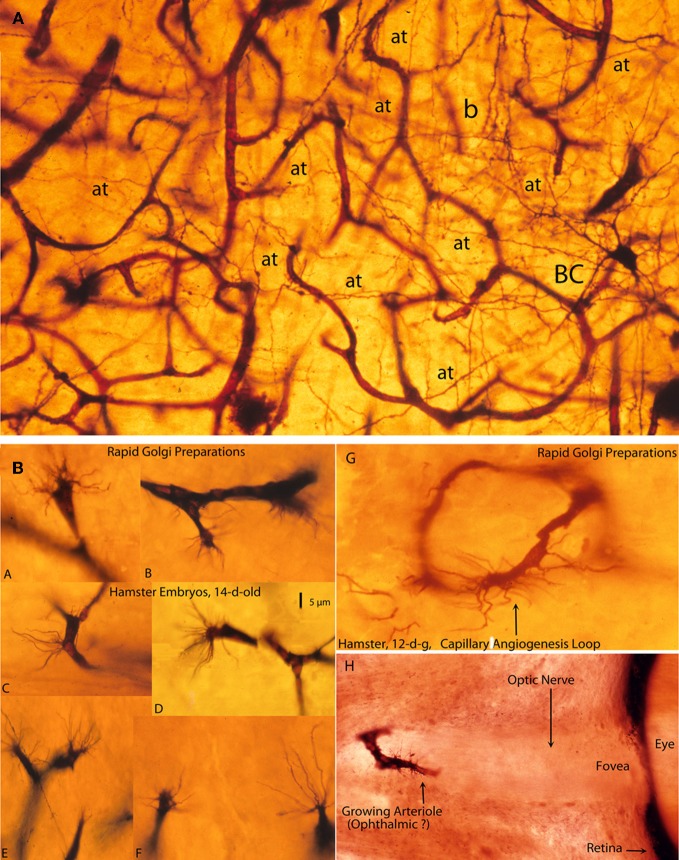
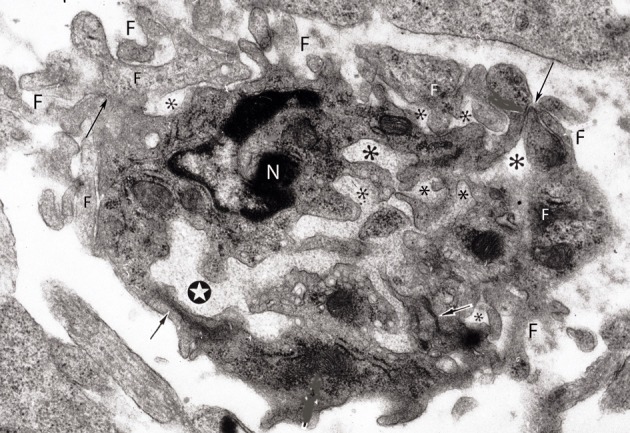
The capillary from the meningeal inner pial lamella play a crucial role in the development and structural organization of the cerebral cortex extrinsic and intrinsic microvascular compartments. Only pial capillaries are capable of perforating through the cortex external glial limiting membrane (EGLM) to enter into the nervous tissue, although incapable of perforating the membrane to exit the brain. Circulatory dynamics and functional demands determine which capillaries become arterial and which capillaries become venous. The perforation of the cortex EGLM by pial capillaries is a complex process characterized by three fundamental stages: (1) pial capillary contact with the EGLM with fusion of vascular and glial basal laminae at the contact site, (2) endothelial cell filopodium penetration through the fussed laminae with the formation of a funnel between them that accompanies it into the nervous tissue while remaining open to the meningeal interstitium and, (3) penetration of the whole capillary carrying the open funnel with it and establishing an extravascular Virchow-Robin Compartment (V-RC) that maintains the perforating vessel extrinsic (outside) the nervous tissue through its entire length. The V-RC is walled internally by the vascular basal lamina and externally by the basal lamina of joined glial cells endfeet. The VRC outer glial wall appear as an extension of the cortex superficial EGLM. All the perforating vessels within the V-RCs constitute the cerebral cortex extrinsic microvascular compartment. These perforating vessels are the only one capable of responding to inflammatory insults. The V-RC remains open (for life) to the meningeal interstitium permitting the exchanges of fluid and of cells between brain and meninges. The V-RC function as the brain sole drainage (prelymphatic) system in both physiological as well as pathological situations. During cortical development, capillaries emerge from the perforating vessels, by endothelial cells growing sprouts analogous to their angiogenesis, entering into their corresponding V-RCs. These new capillaries to enter into the nervous tissue must perforate through the V-RC outer glial wall, a process analogous to the original perforation of the cortex EGLM by pial capillaries. These emerging capillaries are incapable of reentering the V-RCs and/or perforating vessels. As the new capillary enters into the nervous tissue, it becomes surrounded by glial endfeet and carries a single basal lamina (possibly glial). Capillaries emerging from contiguous perforators establish an anastomotic plexus between them, by mechanisms still poorly understood. The capillaries of this anastomotic plexus constitute the cerebral cortex intrinsic microvascular compartment and together constitute the so-called blood-brain-barrier. The intrinsic capillaries are changing and readapting continuously, by both active angiogenesis and reabsorption, to the gray matter neurons developmental and functional needs. The brain intrinsic capillaries are among the most active microvessels of the human body. Unresolved developmental and functional aspects concerning the cerebral cortex intrinsic capillary plexus need to be further investigated.
Keywords: EGLM; endothelial cell filopodium; human brain; intracerebral microvascular system; meningeal inner pial lamella.
Figures
References
-
- Andres K. H. (1976a). Zur Feinstruktur der Arrachnoidal zotten bei Mammalia. Zeitschrift Zellsforschung Mikroskoscopic Anatomy 82, 92–109
-
- Andres K. H. (1967b). Über die Feinstruktur der Arachnoidea und Dura mater von Mammalia. Zeitschrift Zellsforschung Mikroskoscopic Anatomy 82, 272–295 - PubMed
-
- Ausprunk D. H. (1979). Tumor angiogenesis, in Chemical Messenger of the Inflammatory Process, ed Houck J. C. (Amsterdam: North Holland Press; ), 317–351
-
- Bär T., Wolff J. R. (1972). The formation of capillary basal membranes and internal vascularization of the rat's cerebral cortex. Zeitschrift Zellsforschung Mikroskospic Anatomy 133, 231–248 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources