

New concepts in pharmacological efficacy at 7TM receptors: IUPHAR review 2
- PMID: 22994528
- PMCID: PMC3579279
- DOI: 10.1111/j.1476-5381.2012.02223.x
New concepts in pharmacological efficacy at 7TM receptors: IUPHAR review 2
Abstract
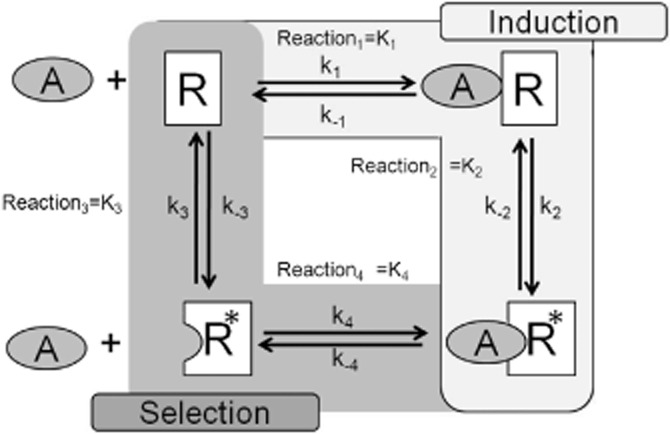
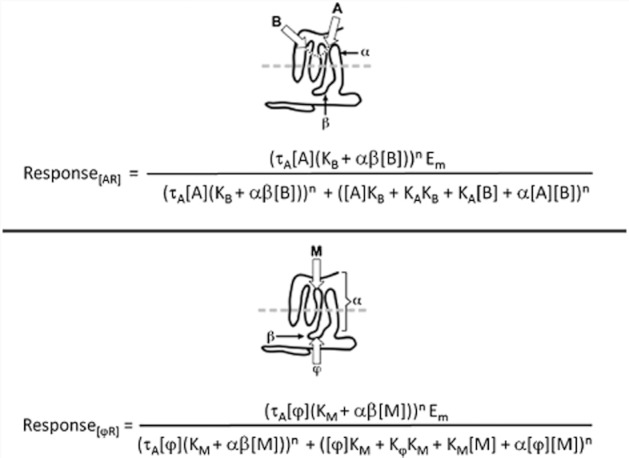
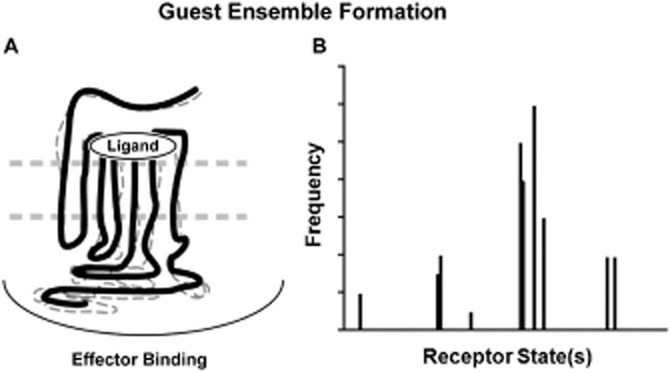
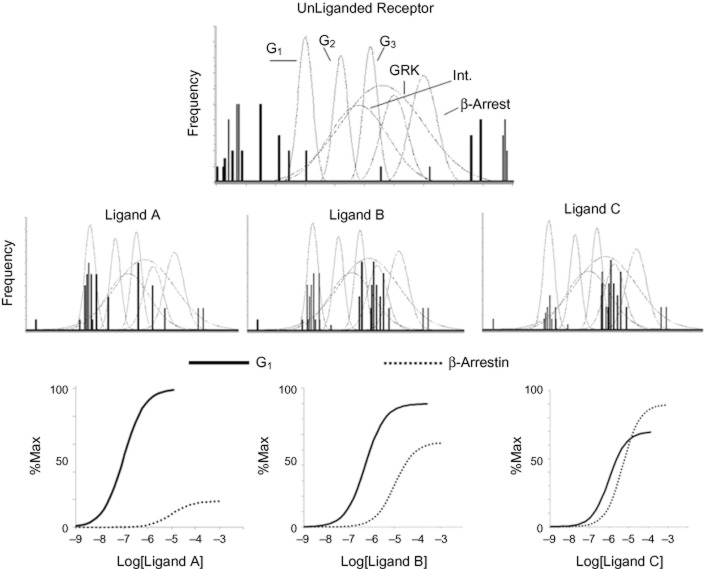
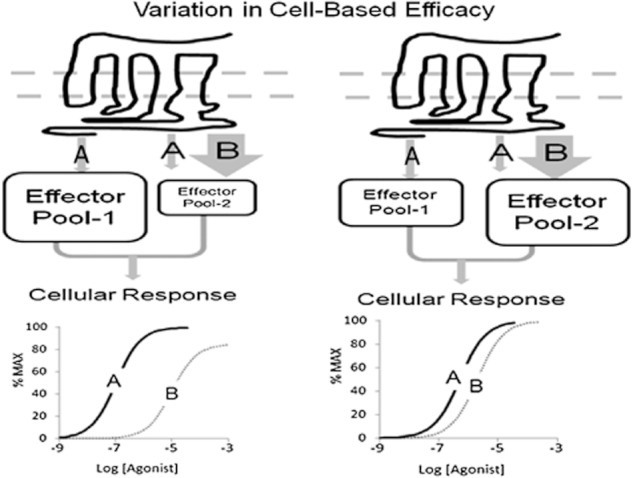
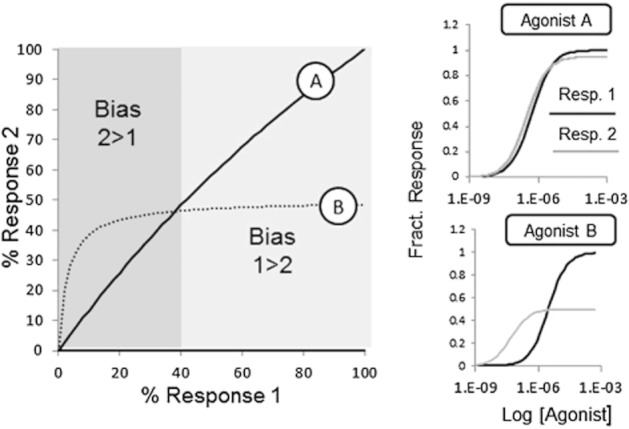
The present-day concept of drug efficacy has changed completely from its original description as the property of agonists that causes tissue activation. The ability to visualize the multiple behaviours of seven transmembrane receptors has shown that drugs can have many efficacies and also that the transduction of drug stimulus to various cellular stimulus-response cascades can be biased towards some but not all pathways. This latter effect leads to agonist 'functional selectivity', which can be favourable for the improvement of agonist therapeutics. However, in addition, biased agonist potency becomes cell type dependent with the loss of the monotonic behaviour of stimulus-response mechanisms, leading to potential problems in agonist quantification. This has an extremely important effect on the discovery process for new agonists since it now cannot be assumed that a given screening or lead optimization assay will correctly predict therapeutic behaviour. This review discusses these ideas and how new approaches to quantifying agonist effect may be used to circumvent the cell type dependence of agonism. This article, written by a corresponding member of the International Union of Basic and Clinical Pharmacology Committee on Receptor Nomenclature and Drug Classification (NC-IUPHAR), reviews our current understanding of the interaction of ligands with seven transmembrane receptors. Further information on these pharmacological concepts is being incorporated into the IUPHAR/BPS database GuideToPharmacology.org.
© 2012 The Author. British Journal of Pharmacology © 2012 The British Pharmacological Society.
Figures


References
-
- AlvarezCurto E, Prihandoko R, Tautermann CS, Zwier JM, Pediani JD, Lohse MJ, et al. Developing chemical genetic approaches to explore G protein-coupled receptor function: validation of the use of a receptor activated solely by synthetic ligand (RASSL) Mol Pharmacol. 2011;80:1033–1046. - PMC - PubMed
-
- Ariens EJ. Affinity and intrinsic activity in the theory of competitive inhibition. Arch Int Pharmacodyn Ther. 1954;99:32–49. - PubMed
-
- Baneres J-L, Mesnier D, Martin A, Joubert L, Dumuis A, Bockaret J. Molecular characterization of a purified 5-HT4 receptor. J Biol Chem. 2005;280:20253–20260. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
