

The crystal structure of the lipid II-degrading bacteriocin syringacin M suggests unexpected evolutionary relationships between colicin M-like bacteriocins
- PMID: 22995910
- PMCID: PMC3493929
- DOI: 10.1074/jbc.M112.400150
The crystal structure of the lipid II-degrading bacteriocin syringacin M suggests unexpected evolutionary relationships between colicin M-like bacteriocins
Abstract
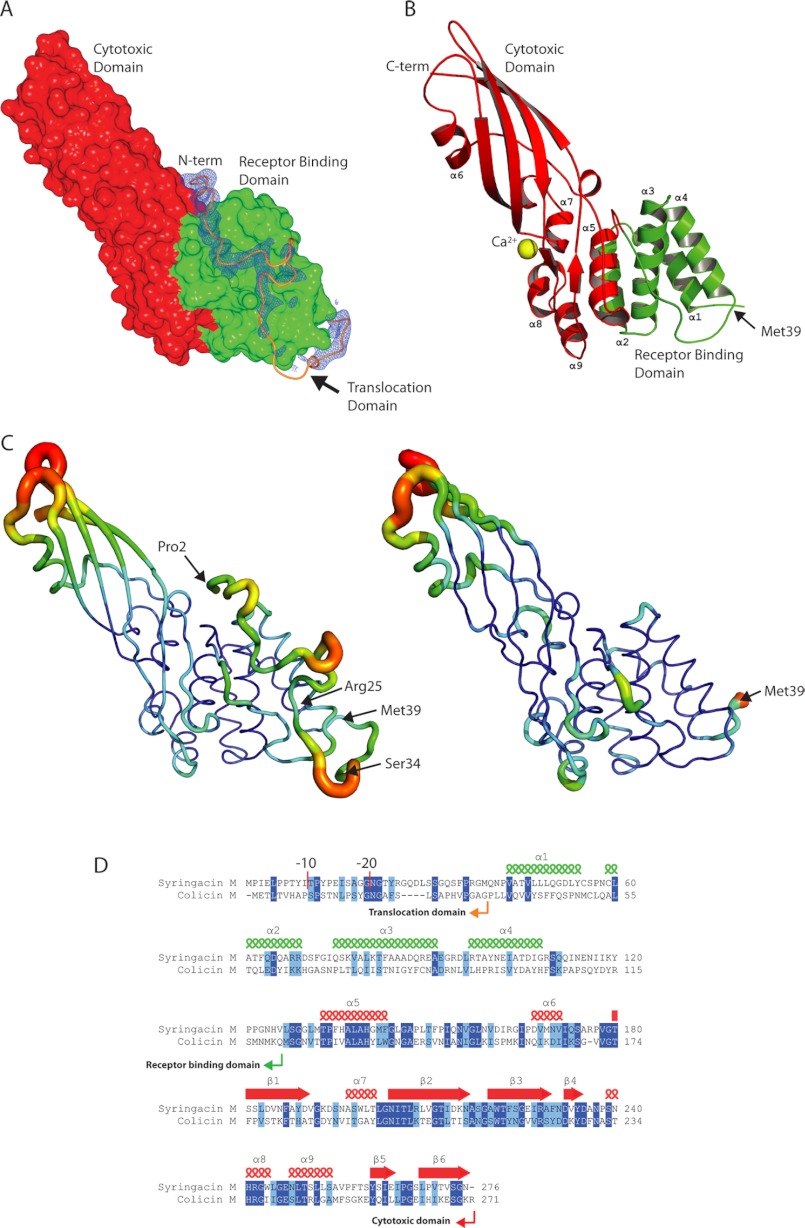
Colicin-like bacteriocins show potential as next generation antibiotics with clinical and agricultural applications. Key to these potential applications is their high potency and species specificity that enables a single pathogenic species to be targeted with minimal disturbance of the wider microbial community. Here we present the structure and function of the colicin M-like bacteriocin, syringacin M from Pseudomonas syringae pv. tomato DC3000. Syringacin M kills susceptible cells through a highly specific phosphatase activity that targets lipid II, ultimately inhibiting peptidoglycan synthesis. Comparison of the structures of syringacin M and colicin M reveals that, in addition to the expected similarity between the homologous C-terminal catalytic domains, the receptor binding domains of these proteins, which share no discernible sequence homology, share a striking structural similarity. This indicates that the generation of the novel receptor binding and species specificities of these bacteriocins has been driven by diversifying selection rather than diversifying recombination as suggested previously. Additionally, the structure of syringacin M reveals the presence of an active site calcium ion that is coordinated by a conserved aspartic acid side chain and is essential for catalytic activity. We show that mutation of this residue to alanine inactivates syringacin M and that the metal ion is absent from the structure of the mutant protein. Consistent with the presence of Ca(2+) in the active site, we show that syringacin M activity is supported by Ca(2+), along with Mg(2+) and Mn(2+), and the protein is catalytically inactive in the absence of these ions.
Figures







Similar articles
-
Functional and structural characterization of PaeM, a colicin M-like bacteriocin produced by Pseudomonas aeruginosa.J Biol Chem. 2012 Oct 26;287(44):37395-405. doi: 10.1074/jbc.M112.406439. Epub 2012 Sep 12. J Biol Chem. 2012. PMID: 22977250 Free PMC article.
-
Human- and plant-pathogenic Pseudomonas species produce bacteriocins exhibiting colicin M-like hydrolase activity towards peptidoglycan precursors.J Bacteriol. 2009 Jun;191(11):3657-64. doi: 10.1128/JB.01824-08. Epub 2009 Apr 3. J Bacteriol. 2009. PMID: 19346308 Free PMC article.
-
Purification and characterization of syringacin 4-A, a bacteriocin from pseudomonas syringae 4-A.Antimicrob Agents Chemother. 1974 Jul;6(1):76-83. doi: 10.1128/AAC.6.1.76. Antimicrob Agents Chemother. 1974. PMID: 15828174 Free PMC article.
-
Characterization of colicin M and its orthologs targeting bacterial cell wall peptidoglycan biosynthesis.Microb Drug Resist. 2012 Jun;18(3):222-9. doi: 10.1089/mdr.2011.0230. Epub 2012 Mar 20. Microb Drug Resist. 2012. PMID: 22432709 Review.
-
Expanding role of lipid II as a target for lantibiotics.Future Microbiol. 2007 Oct;2(5):513-25. doi: 10.2217/17460913.2.5.513. Future Microbiol. 2007. PMID: 17927474 Review.
Cited by
-
Pseudomonas bacteriocin syringacin M released upon desiccation suppresses the growth of sensitive bacteria in plant necrotic lesions.Microb Biotechnol. 2020 Jan;13(1):134-147. doi: 10.1111/1751-7915.13367. Epub 2019 Jan 22. Microb Biotechnol. 2020. PMID: 30672132 Free PMC article.
-
Bacteriocins Targeting Gram-Negative Phytopathogenic Bacteria: Plantibiotics of the Future.Front Microbiol. 2020 Sep 18;11:575981. doi: 10.3389/fmicb.2020.575981. eCollection 2020. Front Microbiol. 2020. PMID: 33042091 Free PMC article. Review.
-
Crystal structure of pectocin M1 reveals diverse conformations and interactions during its initial step via the ferredoxin uptake system.FEBS Open Bio. 2024 Oct;14(10):1731-1745. doi: 10.1002/2211-5463.13874. Epub 2024 Aug 9. FEBS Open Bio. 2024. PMID: 39123319 Free PMC article.
-
Structural determinants for activity and specificity of the bacterial toxin LlpA.PLoS Pathog. 2013 Feb;9(2):e1003199. doi: 10.1371/journal.ppat.1003199. Epub 2013 Feb 28. PLoS Pathog. 2013. PMID: 23468636 Free PMC article.
-
Structure of the bacterial plant-ferredoxin receptor FusA.Nat Commun. 2016 Oct 31;7:13308. doi: 10.1038/ncomms13308. Nat Commun. 2016. PMID: 27796364 Free PMC article.
References
-
- Riley M. A. (1998) Molecular mechanisms of bacteriocin evolution. Annu. Rev. Genet. 32, 255–278 - PubMed
-
- Kleanthous C. (2010) Swimming against the tide. Progress and challenges in our understanding of colicin translocation. Nat. Rev. Microbiol. 8, 843–848 - PubMed
-
- Michel-Briand Y., Baysse C. (2002) The pyocins of Pseudomonas aeruginosa. Biochimie 84, 499–510 - PubMed
-
- Feldgarden M., Riley M. A. (1998) High levels of colicin resistance in Escherichia coli. Evolution 52, 1270–1276 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
