

Outer membrane vesicles of a human commensal mediate immune regulation and disease protection
- PMID: 22999859
- PMCID: PMC3895402
- DOI: 10.1016/j.chom.2012.08.004
Outer membrane vesicles of a human commensal mediate immune regulation and disease protection
Abstract
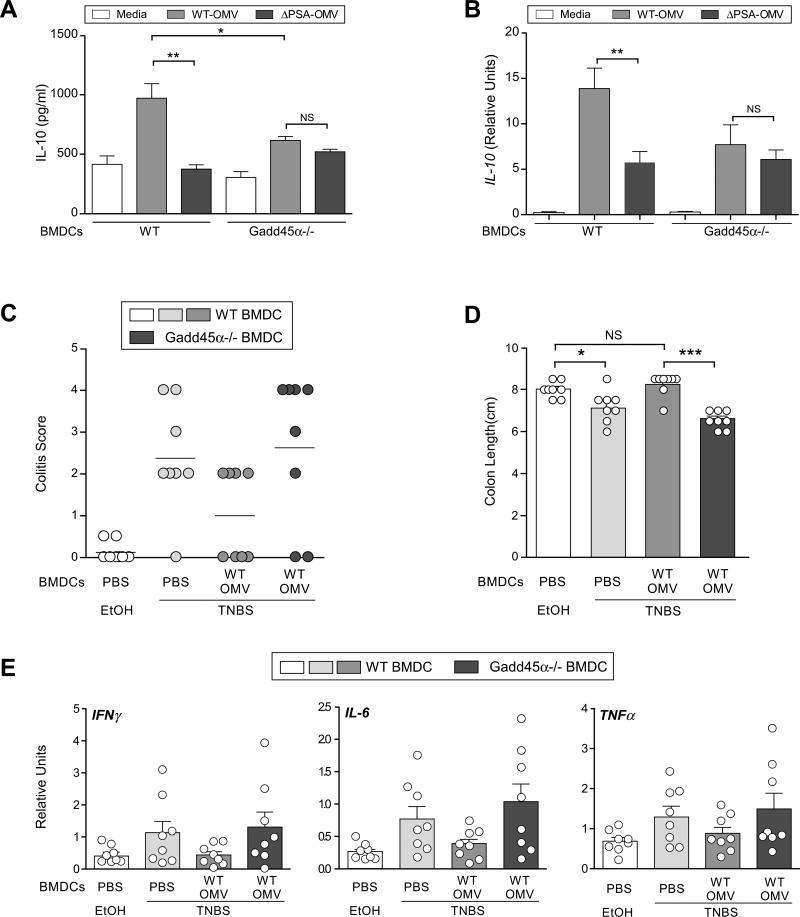
Commensal bacteria impact host health and immunity through various mechanisms, including the production of immunomodulatory molecules. Bacteroides fragilis produces a capsular polysaccharide (PSA), which induces regulatory T cells and mucosal tolerance. However, unlike pathogens, which employ secretion systems, the mechanisms by which commensal bacteria deliver molecules to the host remain unknown. We reveal that Bacteroides fragilis releases PSA in outer membrane vesicles (OMVs) that induce immunomodulatory effects and prevent experimental colitis. Dendritic cells (DCs) sense OMV-associated PSA through TLR2, resulting in enhanced regulatory T cells and anti-inflammatory cytokine production. OMV-induced signaling in DCs requires growth arrest and DNA-damage-inducible protein (Gadd45α). DCs treated with PSA-containing OMVs prevent experimental colitis, whereas Gadd45α(-/-) DCs are unable to promote regulatory T cell responses or suppress proinflammatory cytokine production and host pathology. These findings demonstrate that OMV-mediated delivery of a commensal molecule prevents disease, uncovering a mechanism of interkingdom communication between the microbiota and mammals.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
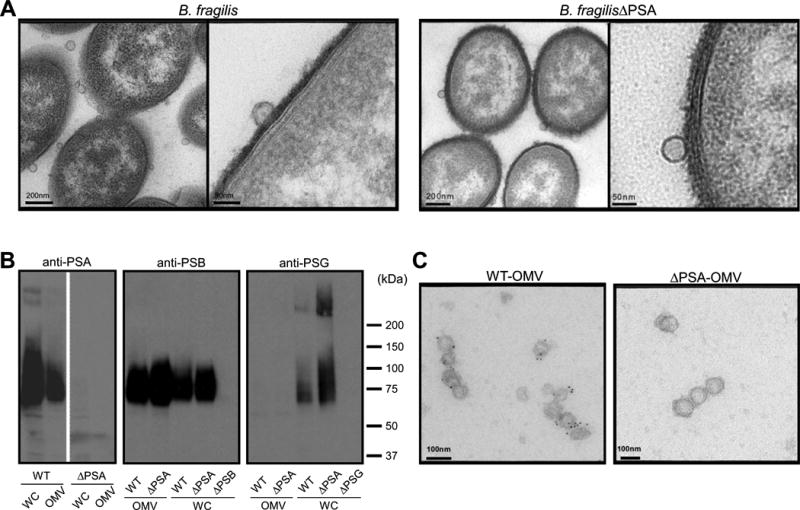
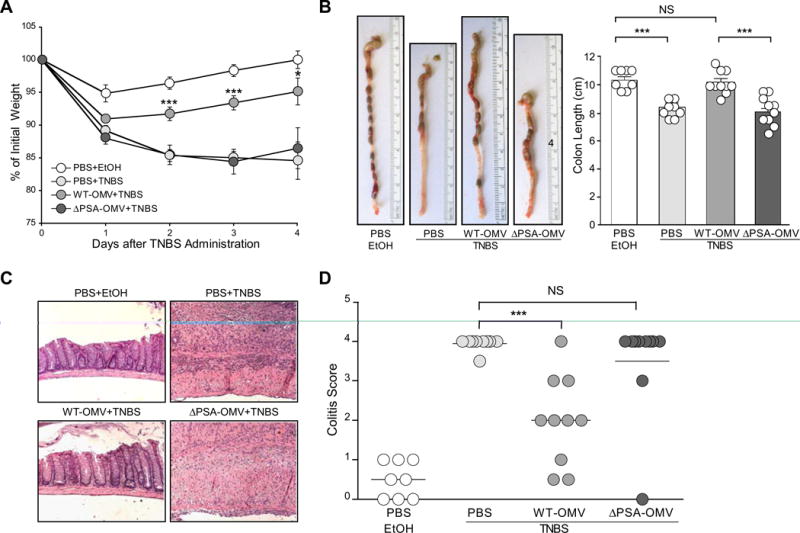
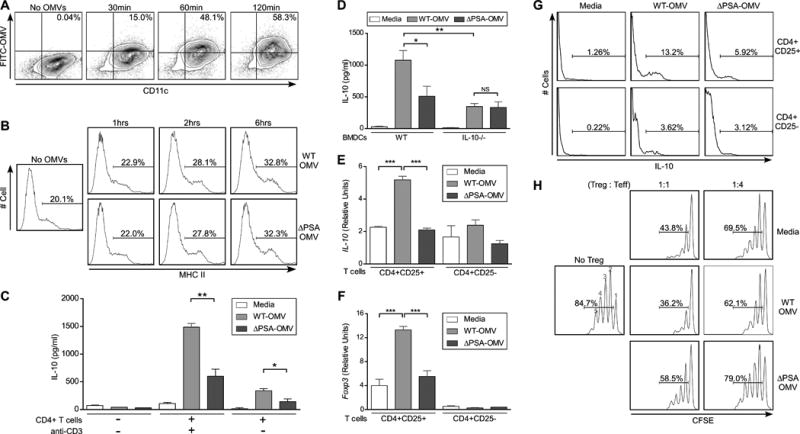
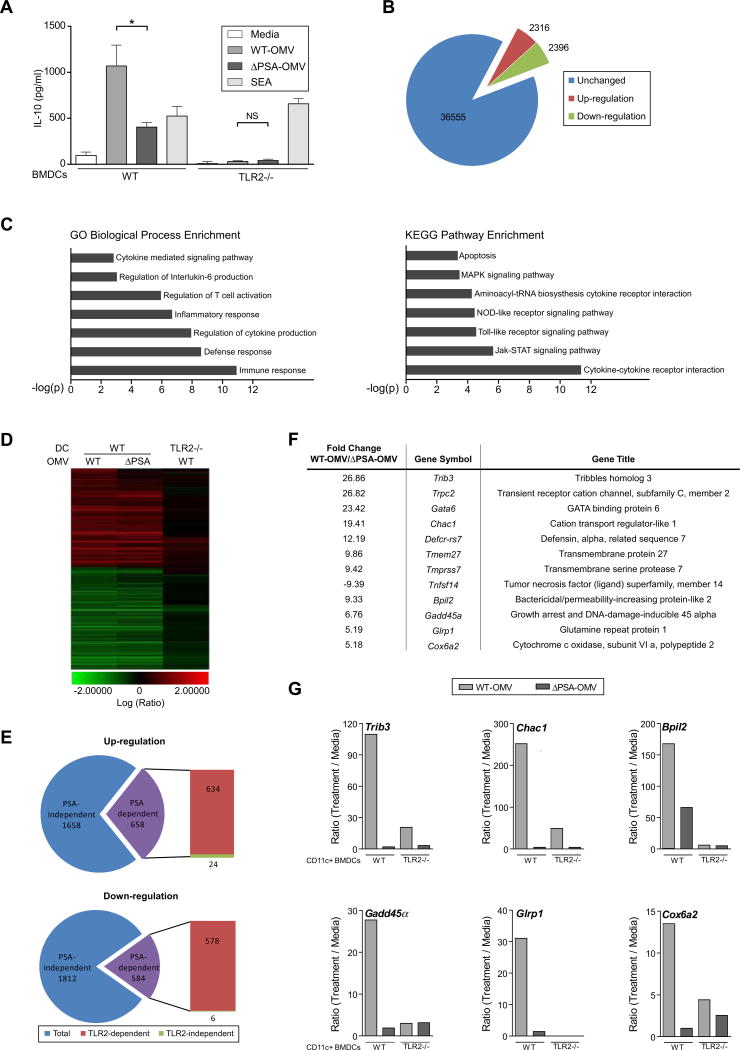
Comment in
-
Secreted bacterial vesicles as good samaritans.Cell Host Microbe. 2012 Oct 18;12(4):392-3. doi: 10.1016/j.chom.2012.10.005. Cell Host Microbe. 2012. PMID: 23084908
References
-
- Cerdeno-Tarraga AM, Patrick S, Crossman LC, Blakely G, Abratt V, Lennard N, Poxton I, Duerden B, Harris B, Quail MA, et al. Extensive DNA inversions in the B. fragilis genome control variable gene expression. Science. 2005;307:1463–1465. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
