

Enthalpic barriers dominate the folding and unfolding of the human Cu, Zn superoxide dismutase monomer
- PMID: 22999954
- PMCID: PMC3712906
- DOI: 10.1016/j.jmb.2012.09.009
Enthalpic barriers dominate the folding and unfolding of the human Cu, Zn superoxide dismutase monomer
Abstract
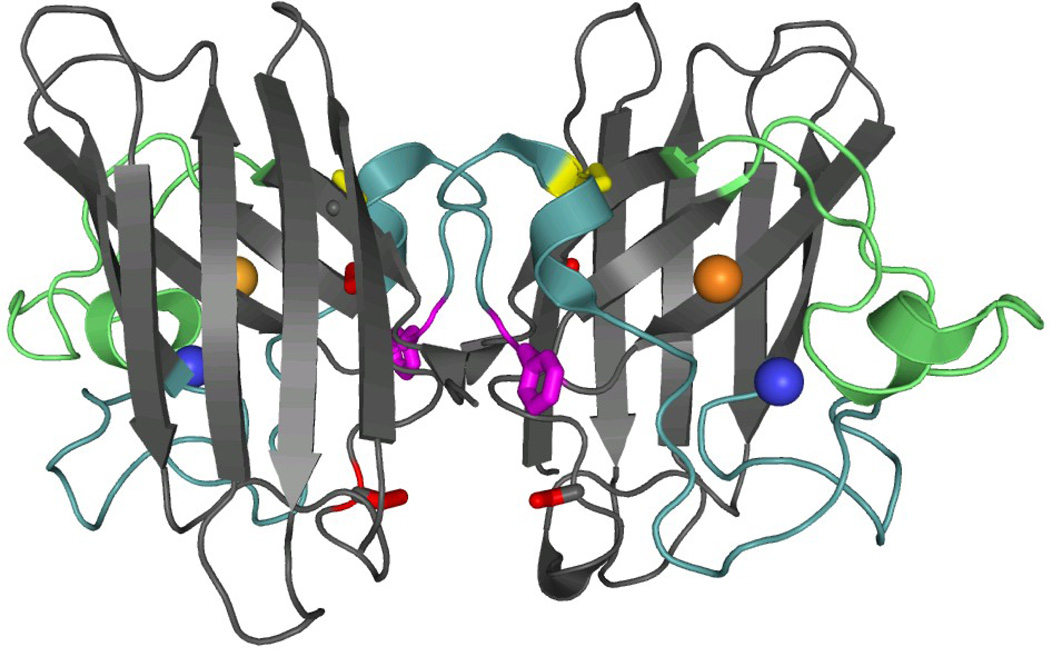
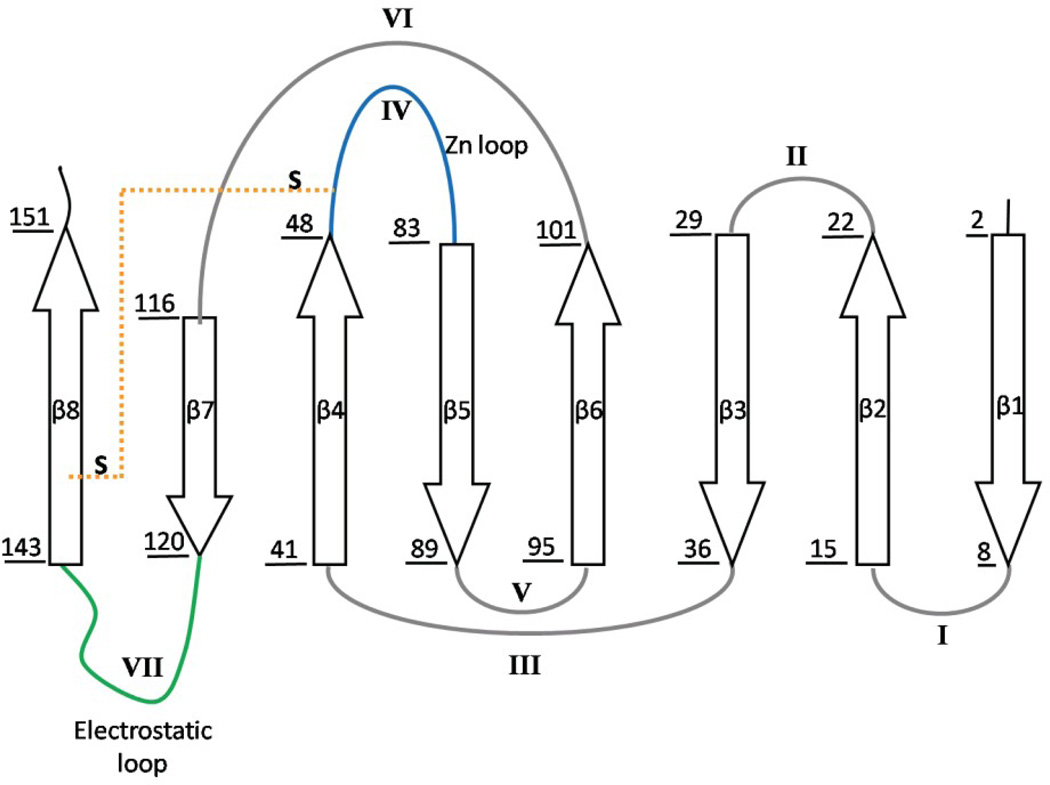
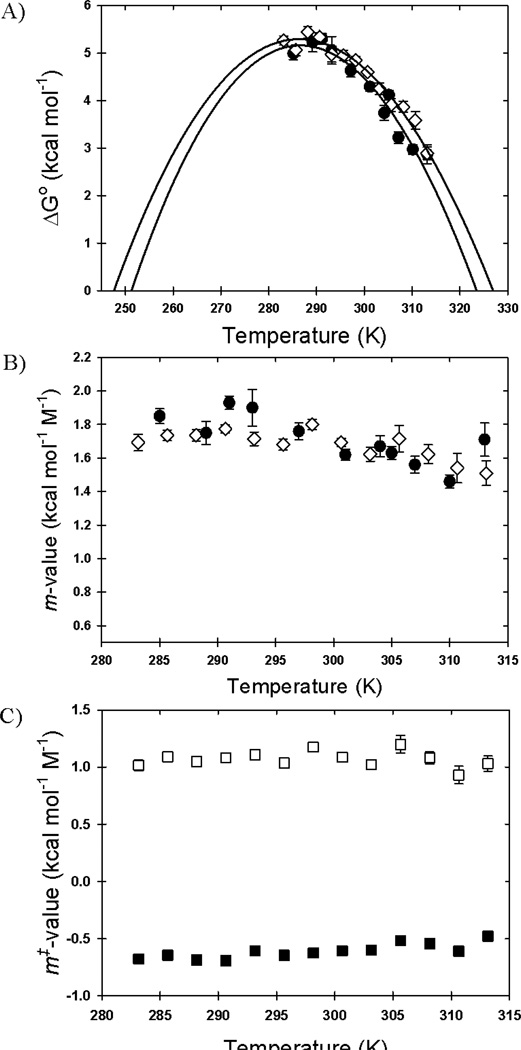
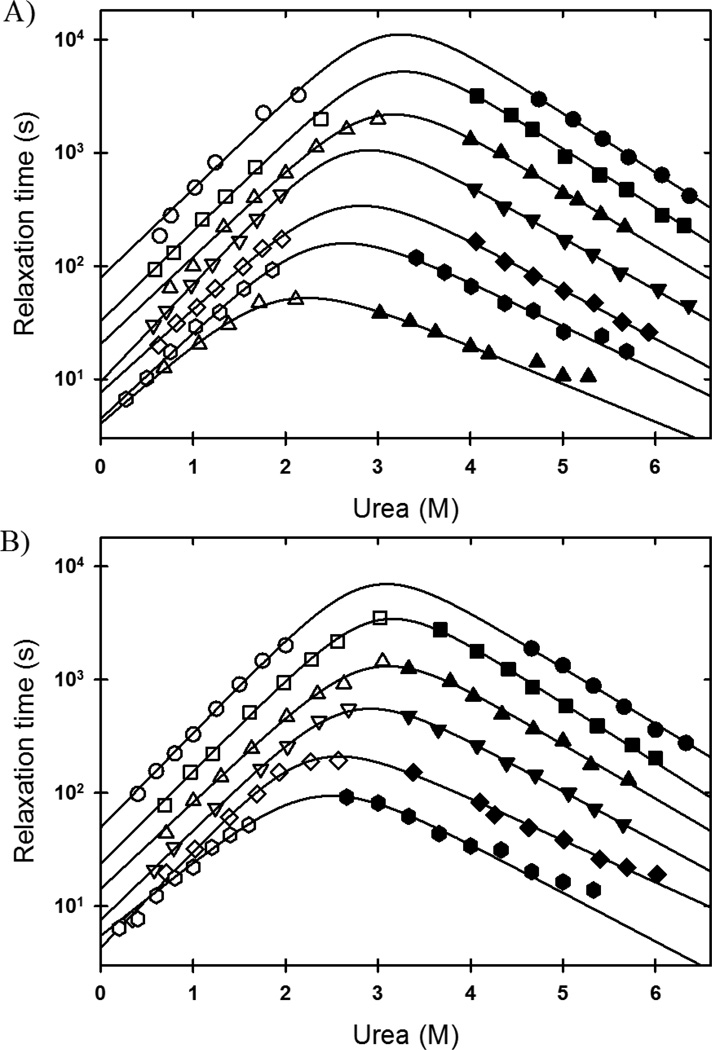
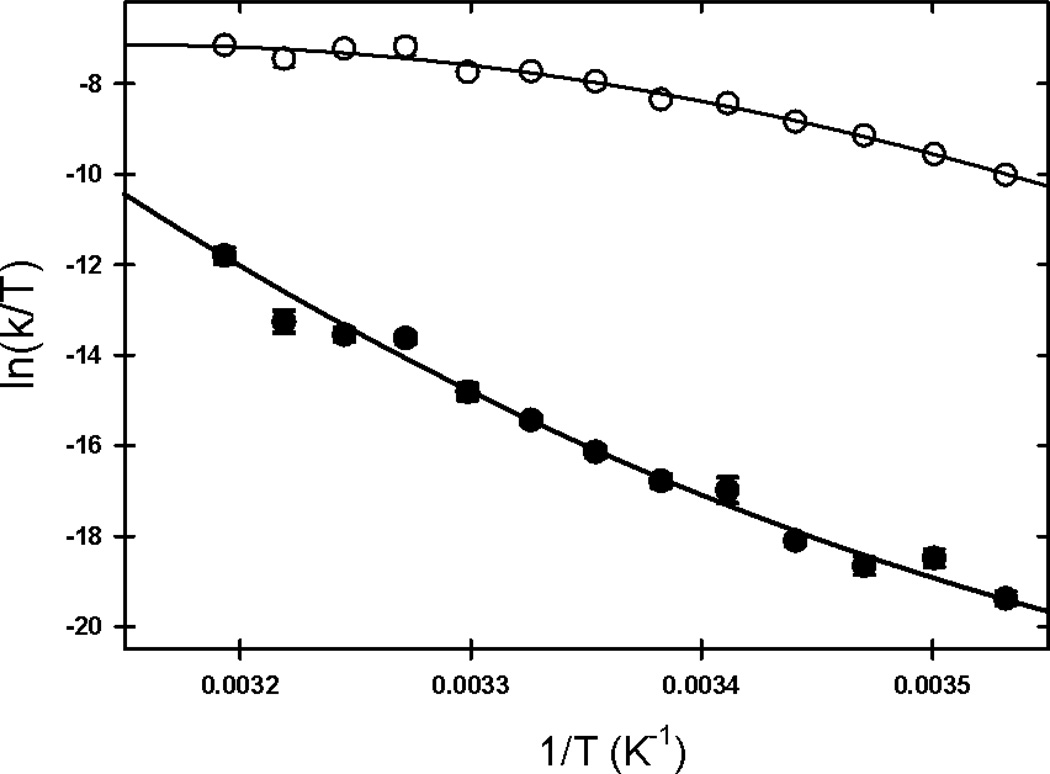
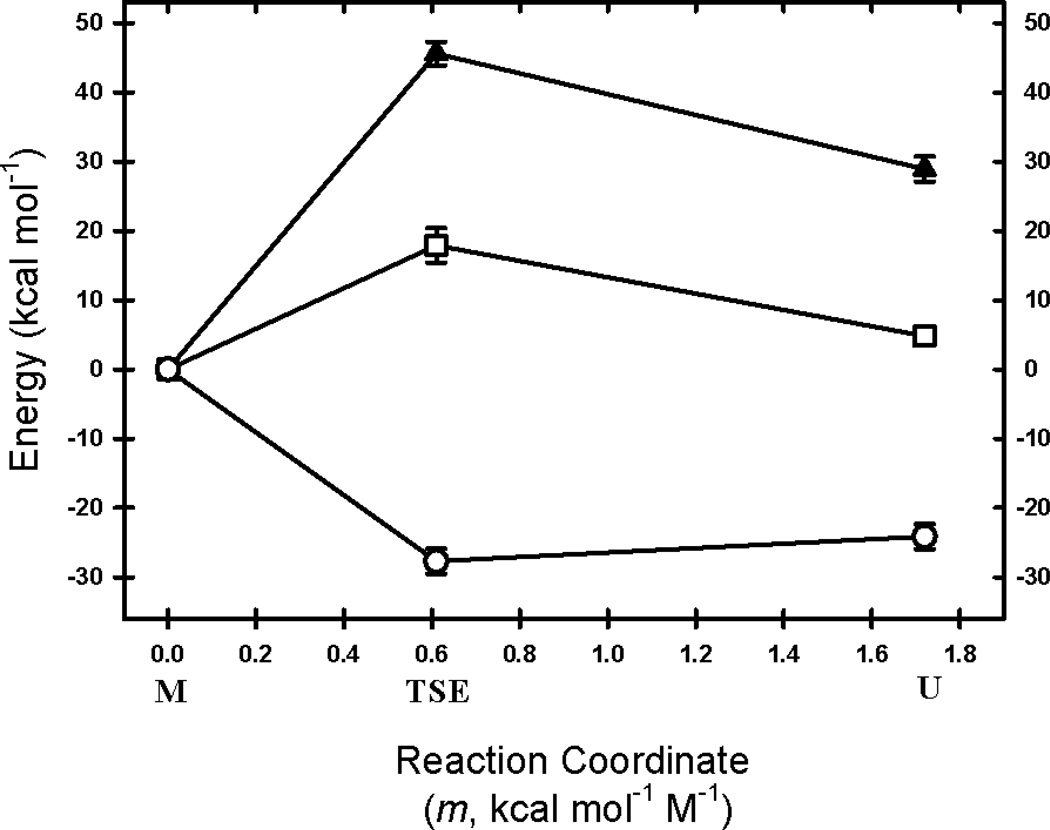
The rate-limiting step in the formation of the native dimeric state of human Cu, Zn superoxide dismutase (SOD1) is a very slow monomer folding reaction that governs the lifetime of its unfolded state. Mutations at dozens of sites in SOD1 are known to cause a fatal motor neuron disease, amyotrophic lateral sclerosis, and recent experiments implicate the unfolded state as a source of soluble oligomers and histologically observable aggregates thought to be responsible for toxicity. To determine the thermodynamic properties of the transition state ensemble (TSE) limiting the folding of this high-contact-order β-sandwich motif, we performed a combined thermal/urea denaturation thermodynamic/kinetic analysis. The barriers to folding and unfolding are dominated by the activation enthalpy at 298 K and neutral pH; the activation entropy is favorable and reduces the barrier height for both reactions. The absence of secondary structure formation or large-scale chain collapse prior to crossing the barrier for folding led to the conclusion that dehydration of nonpolar surfaces in the TSE is responsible for the large and positive activation enthalpy. Although the activation entropy favors the folding reaction, the transition from the unfolded state to the native state is entropically disfavored at 298 K. The opposing entropic contributions to the free energies of the TSE and the native state during folding provide insights into structural properties of the TSE. The results also imply a crucial role for water in governing the productive folding reaction and enhancing the propensity for the aggregation of SOD1.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Zinc binding modulates the entire folding free energy surface of human Cu,Zn superoxide dismutase.J Mol Biol. 2008 Dec 12;384(2):540-55. doi: 10.1016/j.jmb.2008.09.045. Epub 2008 Sep 26. J Mol Biol. 2008. PMID: 18840448 Free PMC article.
-
Disulfide-reduced ALS variants of Cu, Zn superoxide dismutase exhibit increased populations of unfolded species.J Mol Biol. 2010 Apr 30;398(2):320-31. doi: 10.1016/j.jmb.2010.02.034. Epub 2010 Feb 23. J Mol Biol. 2010. PMID: 20184893 Free PMC article.
-
Local unfolding of Cu, Zn superoxide dismutase monomer determines the morphology of fibrillar aggregates.J Mol Biol. 2012 Aug 24;421(4-5):548-60. doi: 10.1016/j.jmb.2011.12.029. Epub 2011 Dec 21. J Mol Biol. 2012. PMID: 22210350 Free PMC article.
-
Structure, folding, and misfolding of Cu,Zn superoxide dismutase in amyotrophic lateral sclerosis.Biochim Biophys Acta. 2006 Nov-Dec;1762(11-12):1025-37. doi: 10.1016/j.bbadis.2006.05.004. Epub 2006 May 22. Biochim Biophys Acta. 2006. PMID: 16814528 Review.
-
Mutant SOD1 instability: implications for toxicity in amyotrophic lateral sclerosis.Neurodegener Dis. 2005;2(3-4):115-27. doi: 10.1159/000089616. Neurodegener Dis. 2005. PMID: 16909016 Review.
Cited by
-
Microsecond barrier-limited chain collapse observed by time-resolved FRET and SAXS.J Mol Biol. 2014 May 1;426(9):1980-94. doi: 10.1016/j.jmb.2014.02.020. Epub 2014 Mar 4. J Mol Biol. 2014. PMID: 24607691 Free PMC article.
-
Nonnative structure in a peptide model of the unfolded state of superoxide dismutase 1 (SOD1): Implications for ALS-linked aggregation.J Biol Chem. 2019 Sep 13;294(37):13708-13717. doi: 10.1074/jbc.RA119.008765. Epub 2019 Jul 24. J Biol Chem. 2019. PMID: 31341015 Free PMC article.
-
Friction-Limited Folding of Disulfide-Reduced Monomeric SOD1.Biophys J. 2020 Apr 21;118(8):1992-2000. doi: 10.1016/j.bpj.2020.02.028. Epub 2020 Mar 12. Biophys J. 2020. PMID: 32191862 Free PMC article.
-
Computational Insights into the Unfolding of a Destabilized Superoxide Dismutase 1 Mutant.Biology (Basel). 2021 Nov 27;10(12):1240. doi: 10.3390/biology10121240. Biology (Basel). 2021. PMID: 34943155 Free PMC article.
-
Cosolutes, Crowding, and Protein Folding Kinetics.J Phys Chem B. 2017 Jul 13;121(27):6527-6537. doi: 10.1021/acs.jpcb.7b03786. Epub 2017 Jun 29. J Phys Chem B. 2017. PMID: 28605189 Free PMC article.
References
-
- Bryngelson JD, Onuchic JN, Socci ND, Wolynes PG. Funnels, pathways, and the energy landscape of protein folding: a synthesis. Proteins. 1995;21:167–195. - PubMed
-
- Plaxco KW, Simons KT, Baker D. Contact order, transition state placement and the refolding rates of single domain proteins. J Mol Biol. 1998;277:985–994. - PubMed
-
- Makhatadze GI, Privalov PL. Energetics of protein structure. Adv Protein Chem. 1995;47:307–425. - PubMed
-
- Becktel WJ, Schellman JA. Protein stability curves. Biopolymers. 1987;26:1859–1877. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
