

A conserved eEF2 coding variant in SCA26 leads to loss of translational fidelity and increased susceptibility to proteostatic insult
- PMID: 23001565
- PMCID: PMC3516132
- DOI: 10.1093/hmg/dds392
A conserved eEF2 coding variant in SCA26 leads to loss of translational fidelity and increased susceptibility to proteostatic insult
Abstract
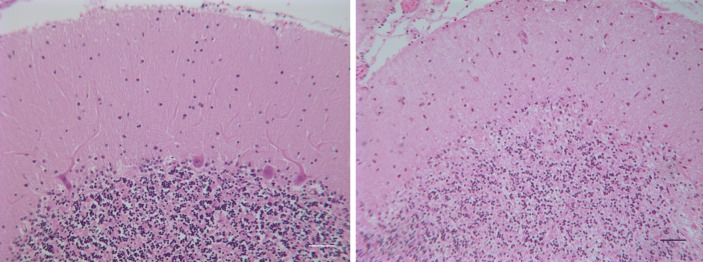
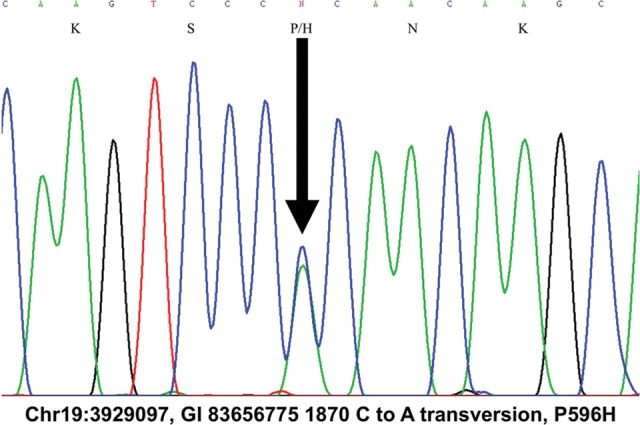
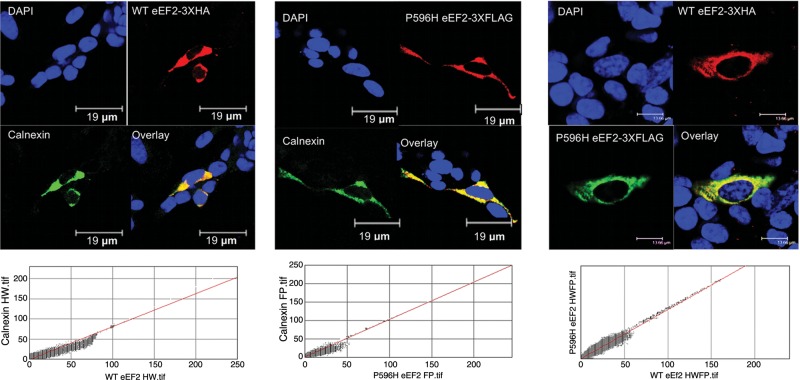
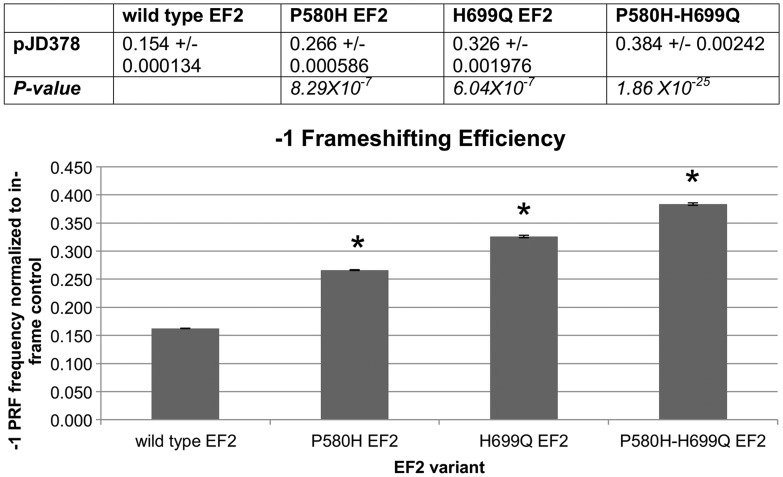
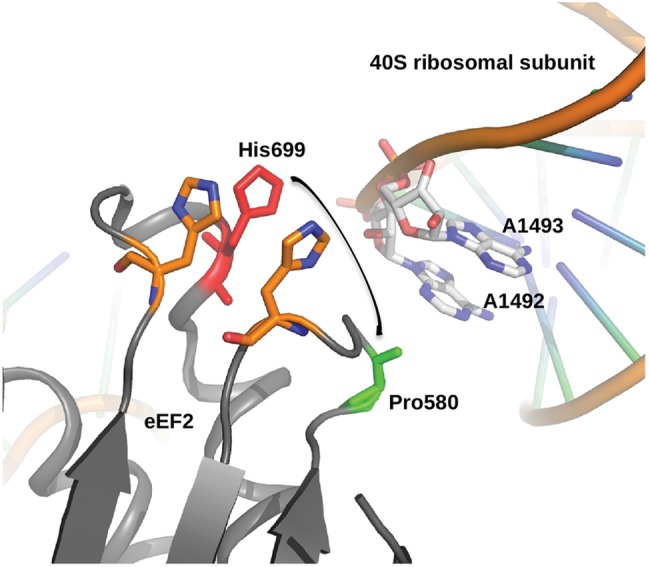
The autosomal dominant spinocerebellar ataxias (SCAs) are a genetically heterogeneous group of disorders exhibiting cerebellar atrophy and Purkinje cell degeneration whose subtypes arise from 31 distinct genetic loci. Our group previously published the locus for SCA26 on chromosome 19p13.3. In this study, we performed targeted deep sequencing of the critical interval in order to identify candidate causative variants in individuals from the SCA26 family. We identified a single variant that co-segregates with the disease phenotype that produces a single amino acid substitution in eukaryotic elongation factor 2. This substitution, P596H, sits in a domain critical for maintaining reading frame during translation. The yeast equivalent, P580H EF2, demonstrated impaired translocation, detected as an increased rate of -1 programmed ribosomal frameshift read-through in a dual-luciferase assay for observing translational recoding. This substitution also results in a greater susceptibility to proteostatic disruption, as evidenced by a more robust activation of a reporter gene driven by unfolded protein response activation upon challenge with dithiothreitol or heat shock in our yeast model system. Our results present a compelling candidate mutation and mechanism for the pathogenesis of SCA26 and further support the role of proteostatic disruption in neurodegenerative diseases.
Figures


References
-
- Craig K., Keers S.M., Archibald K., Curtis A., Chinnery P.F. Molecular epidemiology of spinocerebellar ataxia type 6. Ann. Neurol. 2004;55:752–755. doi:10.1002/ana.20110. - DOI - PubMed
-
- Manto M.U. The wide spectrum of spinocerebellar ataxias (SCAs) Cerebellum. 2005;4:2–6. doi:10.1080/14734220510007914. - DOI - PubMed
-
- Zoghbi H.Y., Orr H.T. Pathogenic mechanisms of a polyglutamine-mediated neurodegenerative disease, spinocerebellar ataxia type 1. J. Biol. Chem. 2009;284:7425–7429. doi:10.1074/jbc.R800041200. - DOI - PMC - PubMed
-
- Kang S., Hong S. Molecular pathogenesis of spinocerebellar ataxia type 1 disease. Mol. Cells. 2009;27:621–627. doi:10.1007/s10059-009-0095-y. - DOI - PubMed
-
- Lastres-Becker I., Rub U., Auburger G. Spinocerebellar ataxia 2 (SCA2) Cerebellum. 2008;7:115–124. doi:10.1007/s12311-008-0019-y. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
