

An inherited TUBB2B mutation alters a kinesin-binding site and causes polymicrogyria, CFEOM and axon dysinnervation
- PMID: 23001566
- PMCID: PMC3516133
- DOI: 10.1093/hmg/dds393
An inherited TUBB2B mutation alters a kinesin-binding site and causes polymicrogyria, CFEOM and axon dysinnervation
Abstract
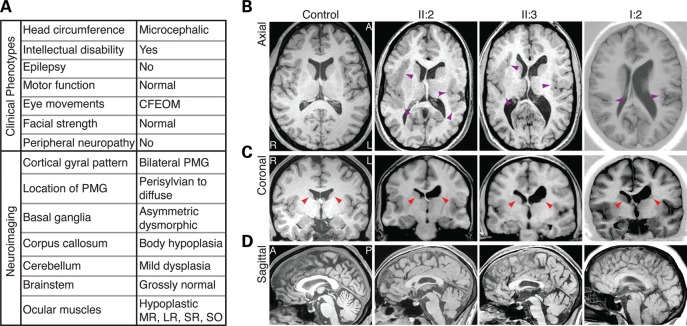
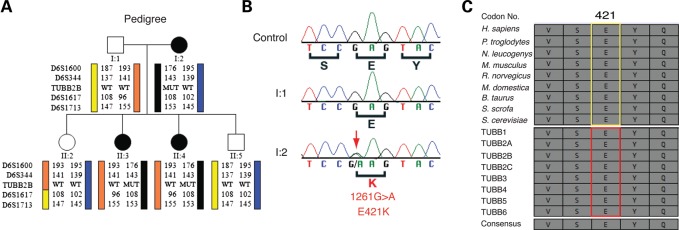
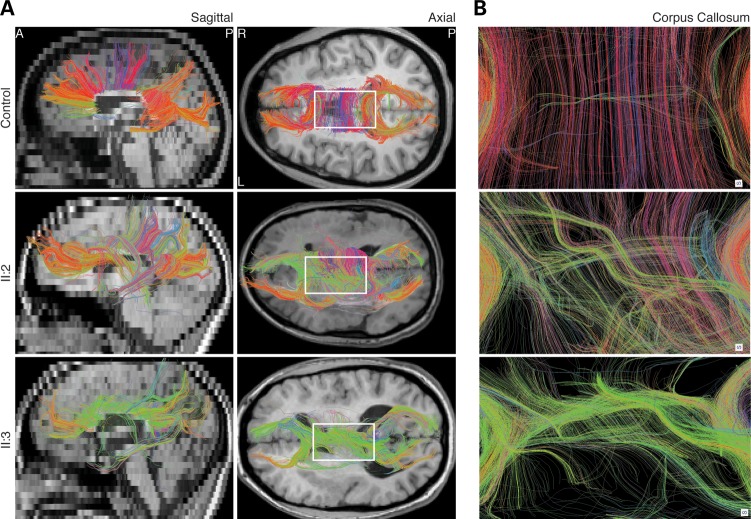
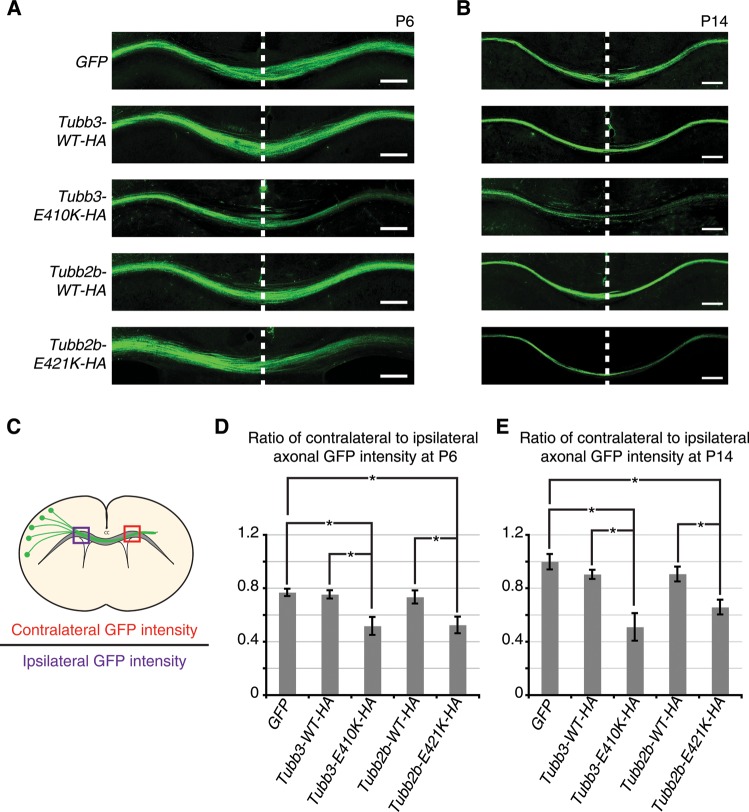
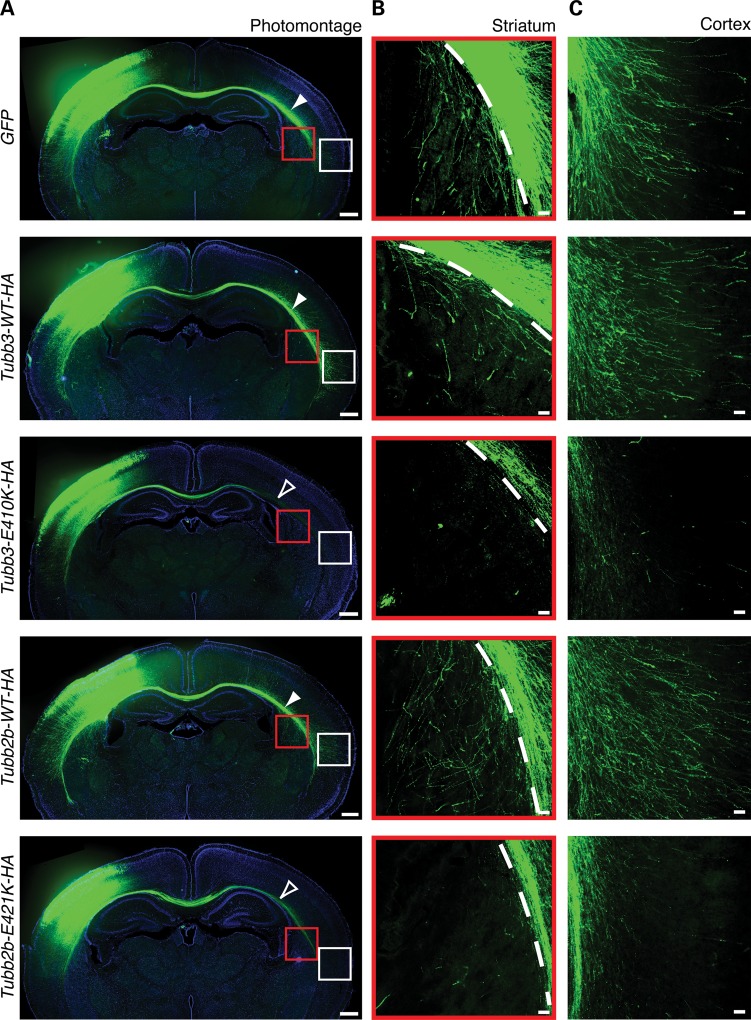
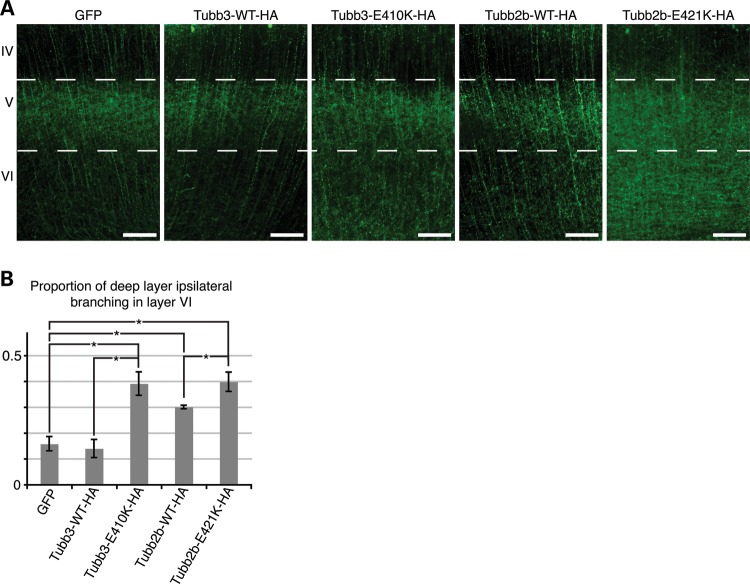
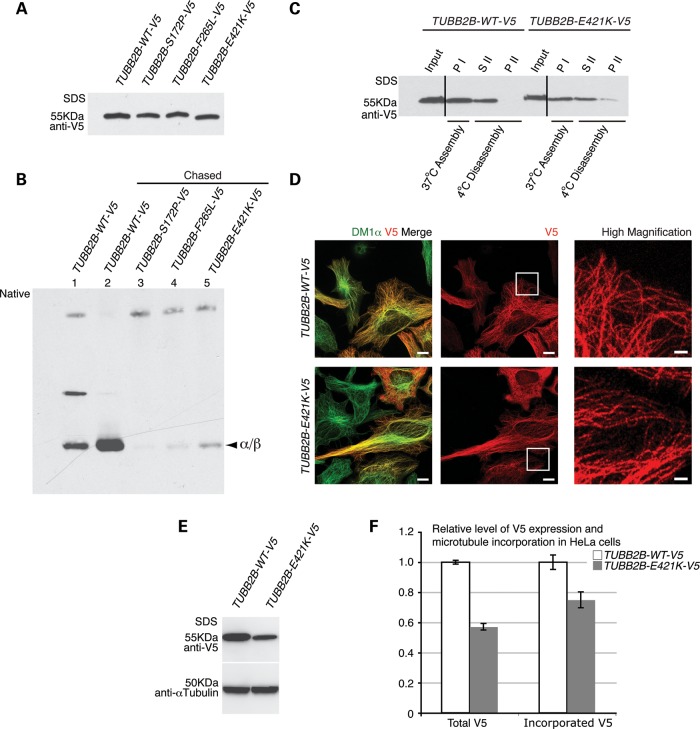
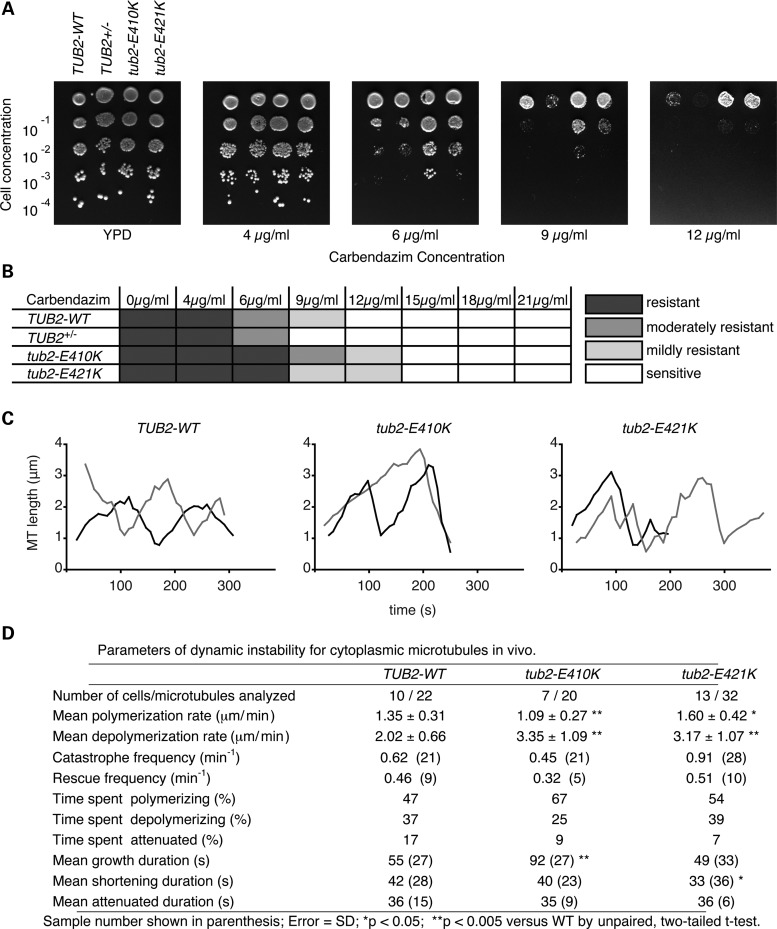
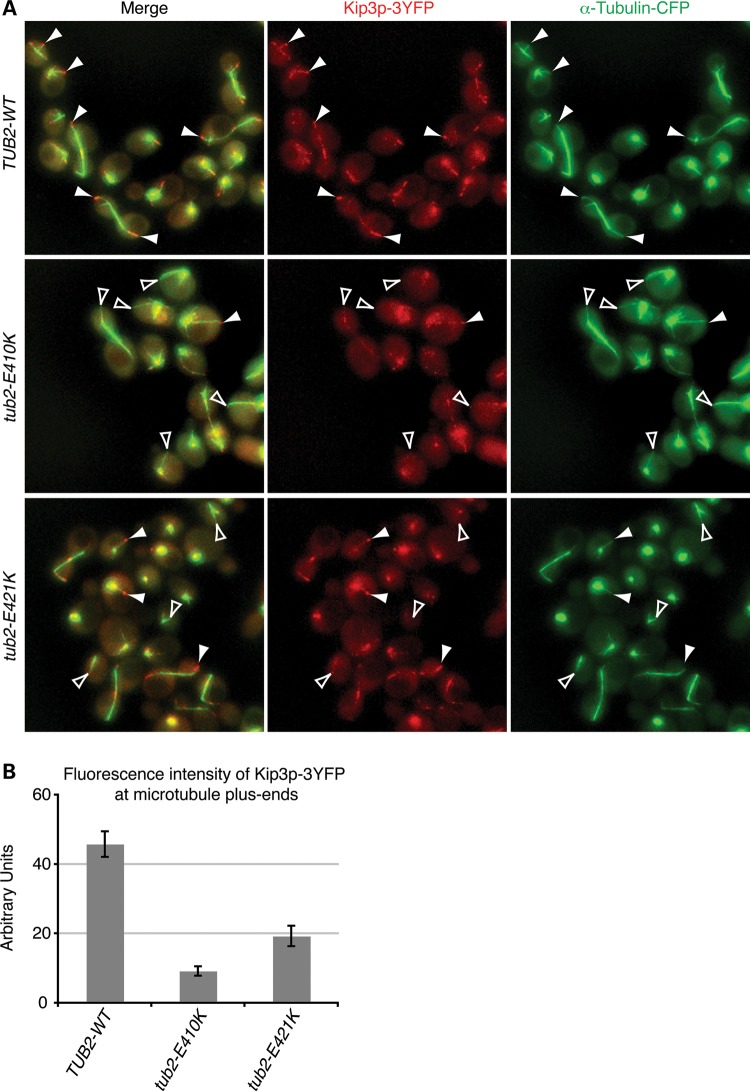
Microtubules are essential components of axon guidance machinery. Among β-tubulin mutations, only those in TUBB3 have been shown to cause primary errors in axon guidance. All identified mutations in TUBB2B result in polymicrogyria, but it remains unclear whether TUBB2B mutations can cause axon dysinnervation as a primary phenotype. We have identified a novel inherited heterozygous missense mutation in TUBB2B that results in an E421K amino acid substitution in a family who segregates congenital fibrosis of the extraocular muscles (CFEOM) with polymicrogyria. Diffusion tensor imaging of brains of affected family members reveals aberrations in the trajectories of commissural projection neurons, implying a paucity of homotopic connections. These observations led us to ask whether axon dysinnervation is a primary phenotype, and why the E421K, but not other, TUBB2B substitutions cause CFEOM. Expression of exogenous Tubb2b-E421K in developing callosal projection neurons is sufficient to perturb homotopic connectivity, without affecting neuronal production or migration. Using in vitro biochemical assays and yeast genetics, we find that TUBB2B-E421K αβ-heterodimers are incorporated into the microtubule network where they alter microtubule dynamics and can reduce kinesin localization. These data provide evidence that TUBB2B mutations can cause primary axon dysinnervation. Interestingly, by incorporating into microtubules and altering their dynamic properties, the E421K substitution behaves differently than previously identified TUBB2B substitutions, providing mechanistic insight into the divergence between resulting phenotypes. Together with previous studies, these findings highlight that β-tubulin isotypes function in both conserved and divergent ways to support proper human nervous system development.
Figures
References
-
- Little M., Seehaus T. Comparative analysis of tubulin sequences. Comp. Biochem. Physiol. B. 1988;90:655–670. - PubMed
-
- Kaltschmidt B., Glatzer K.H., Michiels F., Leiss D., Renkawitz-Pohl R. During Drosophila spermatogenesis beta 1, beta 2 and beta 3 tubulin isotypes are cell-type specifically expressed but have the potential to coassemble into the axoneme of transgenic flies. Eur. J. Cell. Biol. 1991;54:110–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
