

Selection for low dormancy in annual ryegrass (Lolium rigidum) seeds results in high constitutive expression of a glucose-responsive α-amylase isoform
- PMID: 23002268
- PMCID: PMC3503496
- DOI: 10.1093/aob/mcs213
Selection for low dormancy in annual ryegrass (Lolium rigidum) seeds results in high constitutive expression of a glucose-responsive α-amylase isoform
Abstract
Background and aims: α-Amylase in grass caryopses (seeds) is usually expressed upon commencement of germination and is rarely seen in dry, mature seeds. A heat-stable α-amylase activity was unexpectedly selected for expression in dry annual ryegrass (Lolium rigidum) seeds during targeted selection for low primary dormancy. The aim of this study was to characterize this constitutive activity biochemically and determine if its presence conferred insensitivity to the germination inhibitors abscisic acid and benzoxazolinone.
Methods: α-Amylase activity in developing, mature and germinating seeds from the selected (low-dormancy) and a field-collected (dormant) population was characterized by native activity PAGE. The response of seed germination and α-amylase activity to abscisic acid and benzoxazolinone was assessed. Using an alginate affinity matrix, α-amylase was purified from dry and germinating seeds for analysis of its enzymatic properties.
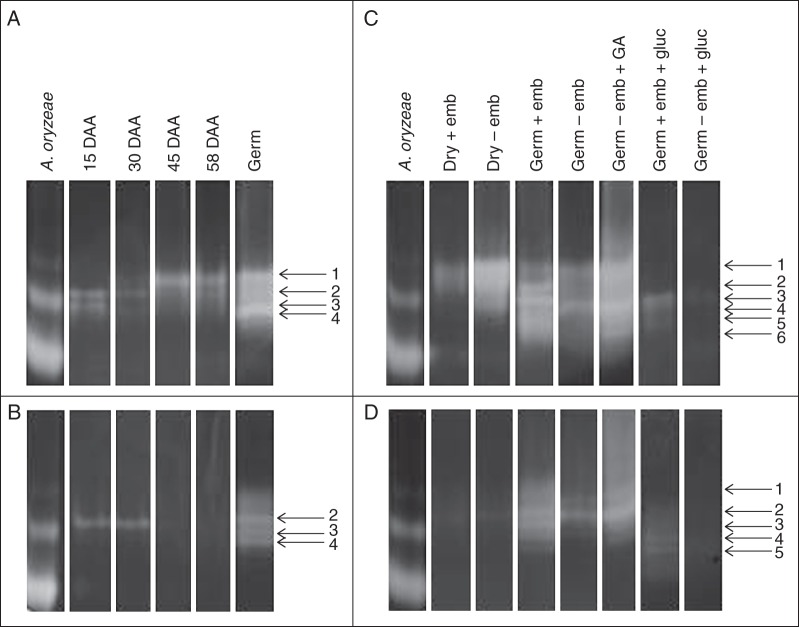
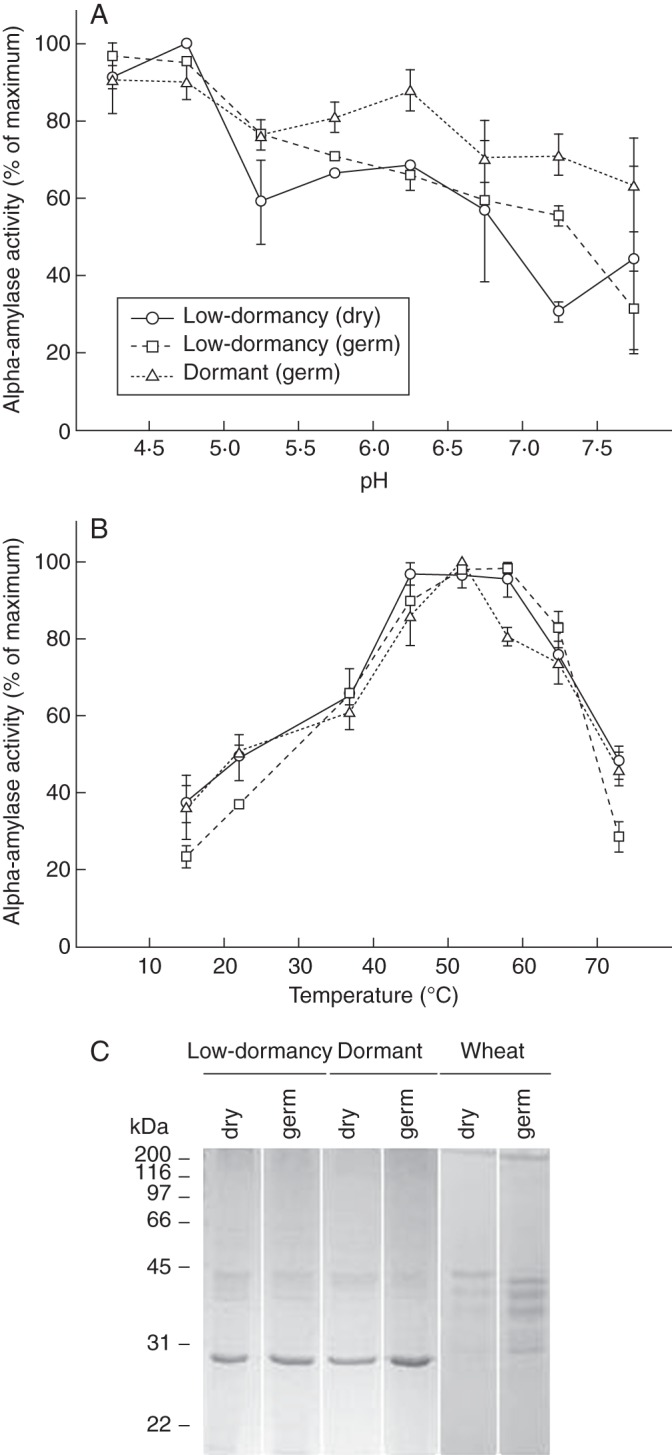
Key results: The constitutive α-amylase activity appeared late during seed development and was mainly localized in the aleurone; in germinating seeds, this activity was responsive to both glucose and gibberellin. It migrated differently on native PAGE compared with the major activities in germinating seeds of the dormant population, but the enzymatic properties of α-amylase purified from the low-dormancy and dormant seeds were largely indistinguishable. Seed imbibition on benzoxazolinone had little effect on the low-dormancy seeds but greatly inhibited germination and α-amylase activity in the dormant population.
Conclusions: The constitutive α-amylase activity in annual ryegrass seeds selected for low dormancy is electrophoretically different from that in germinating seeds and its presence confers insensitivity to benzoxazolinone. The concurrent selection of low dormancy and constitutive α-amylase activity may help to enhance seedling establishment under competitive conditions.
Figures


References
-
- Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T. Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant and Cell Physiology. 2005;46:858–869. - PubMed
-
- Beck E, Ziegler P. Biosynthesis and degradation of starch in higher plants. Annual Review of Plant Physiology and Plant Molecular Biology. 1989;40:95–117.
-
- Bewley JD, Black M. Seeds: physiology of development and germination. 2nd edn. New York: Plenum Press; 1994.
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Analytical Biochemistry. 1976;72:248–254. - PubMed
-
- Candiano G, Bruschi M, Musante L, et al. Blue-silver: a very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis. 2004;25:1327–1333. - PubMed
