

Glucose modulates respiratory complex I activity in response to acute mitochondrial dysfunction
- PMID: 23007390
- PMCID: PMC3493916
- DOI: 10.1074/jbc.M112.386060
Glucose modulates respiratory complex I activity in response to acute mitochondrial dysfunction
Abstract
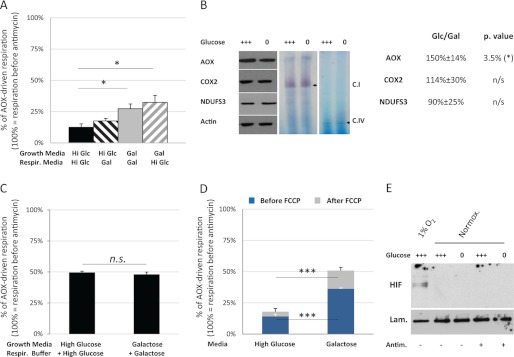
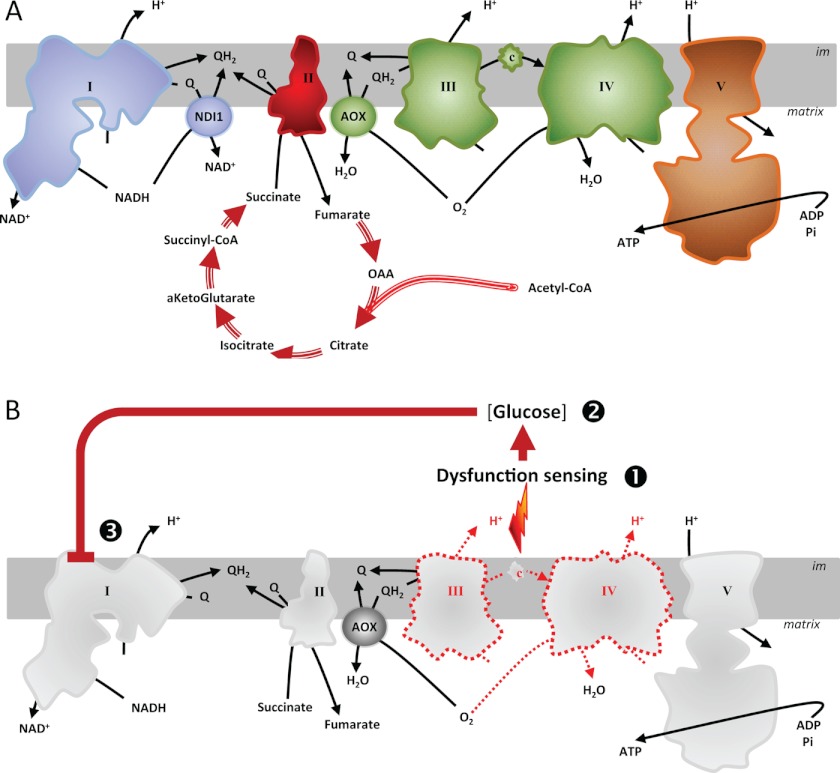
Proper coordination between glycolysis and respiration is essential, yet the regulatory mechanisms involved in sensing respiratory chain defects and modifying mitochondrial functions accordingly are unclear. To investigate the nature of this regulation, we introduced respiratory bypass enzymes into cultured human (HEK293T) cells and studied mitochondrial responses to respiratory chain inhibition. In the absence of respiratory chain inhibitors, the expression of alternative respiratory enzymes did not detectably alter cell physiology or mitochondrial function. However, in permeabilized cells NDI1 (alternative NADH dehydrogenase) bypassed complex I inhibition, whereas alternative oxidase (AOX) bypassed complex III or IV inhibition. In contrast, in intact cells the effects of the AOX bypass were suppressed by growth on glucose, whereas those produced by NDI1 were unaffected. Moreover, NDI1 abolished the glucose suppression of AOX-driven respiration, implicating complex I as the target of this regulation. Rapid Complex I down-regulation was partly released upon prolonged respiratory inhibition, suggesting that it provides an "emergency shutdown" system to regulate metabolism in response to dysfunctions of the oxidative phosphorylation. This system was independent of HIF1, mitochondrial superoxide, or ATP synthase regulation. Our findings reveal a novel pathway for adaptation to mitochondrial dysfunction and could provide new opportunities for combatting diseases.
Figures
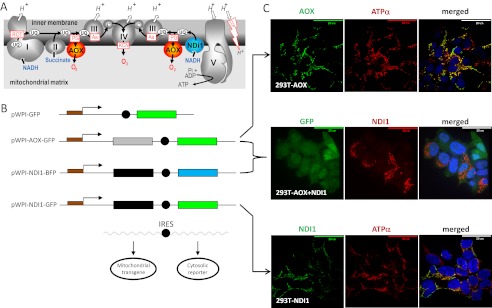
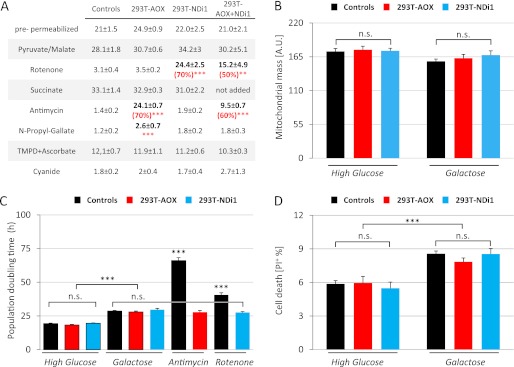
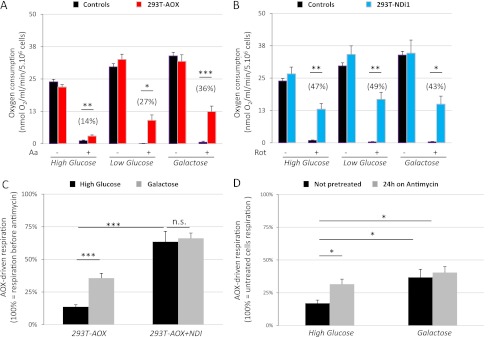
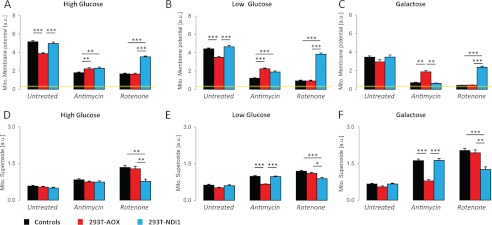
References
-
- DiMauro S. (2004) Mitochondrial diseases. Biochim. Biophys. Acta 1658, 80–88 - PubMed
-
- Zeviani M., Carelli V. (2007) Mitochondrial disorders. Curr. Opin. Neurol. 20, 564–571 - PubMed
-
- DiMauro S., Schon E. A. (2008) Mitochondrial disorders in the nervous system. Annu. Rev. Neurosci. 31, 91–123 - PubMed
-
- Marroquin L. D., Hynes J., Dykens J. A., Jamieson J. D., Will Y. (2007) Circumventing the Crabtree effect. Replacing media glucose with galactose increases susceptibility of HepG2 cells to mitochondrial toxicants. Toxicol. Sci. 97, 539–547 - PubMed
-
- Díaz-Ruiz R., Avéret N., Araiza D., Pinson B., Uribe-Carvajal S., Devin A., Rigoulet M. (2008) Mitochondrial oxidative phosphorylation is regulated by fructose 1,6-bisphosphate. A possible role in Crabtree effect induction? J. Biol. Chem. 283, 26948–26955 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
