

An amino acid substitution inhibits specialist herbivore production of an antagonist effector and recovers insect-induced plant defenses
- PMID: 23008466
- PMCID: PMC3490598
- DOI: 10.1104/pp.112.201061
An amino acid substitution inhibits specialist herbivore production of an antagonist effector and recovers insect-induced plant defenses
Abstract
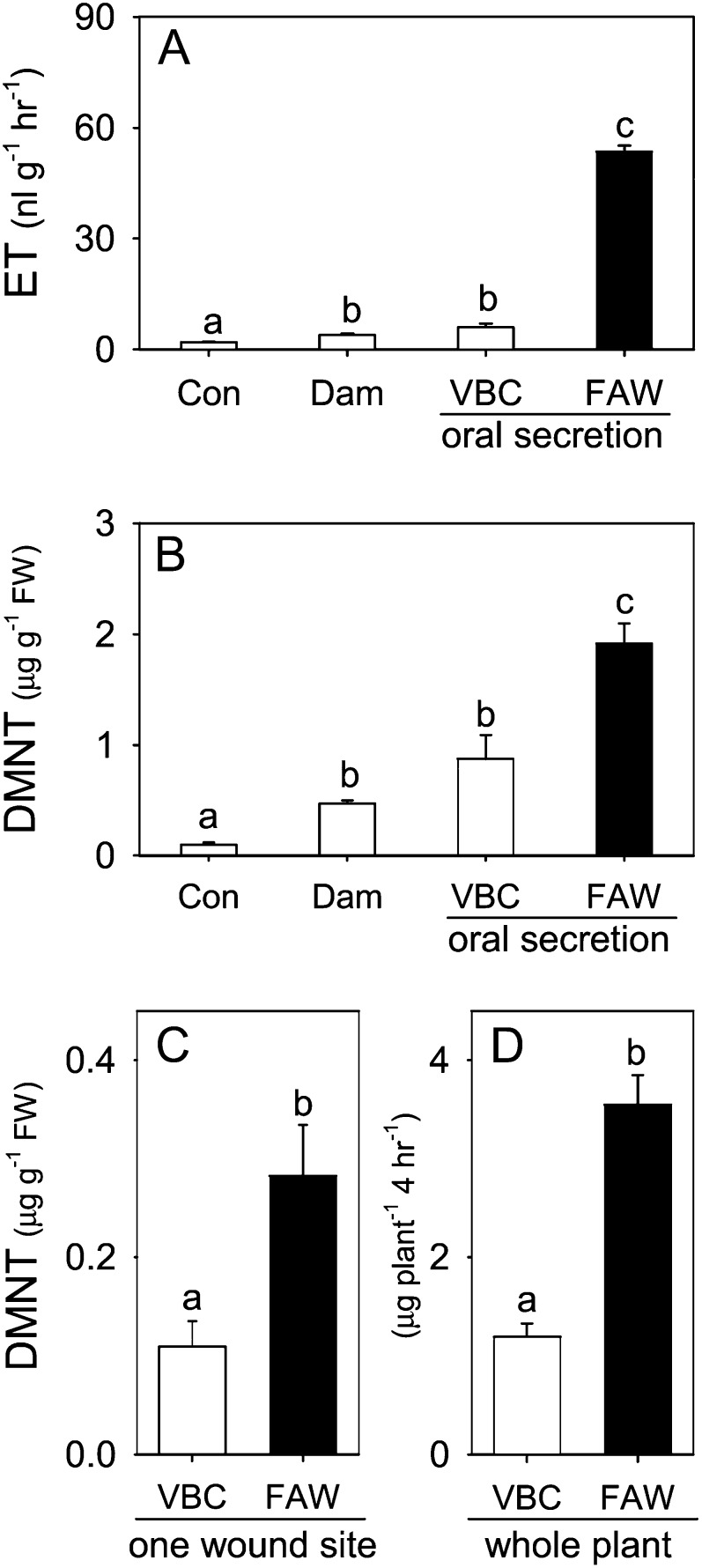
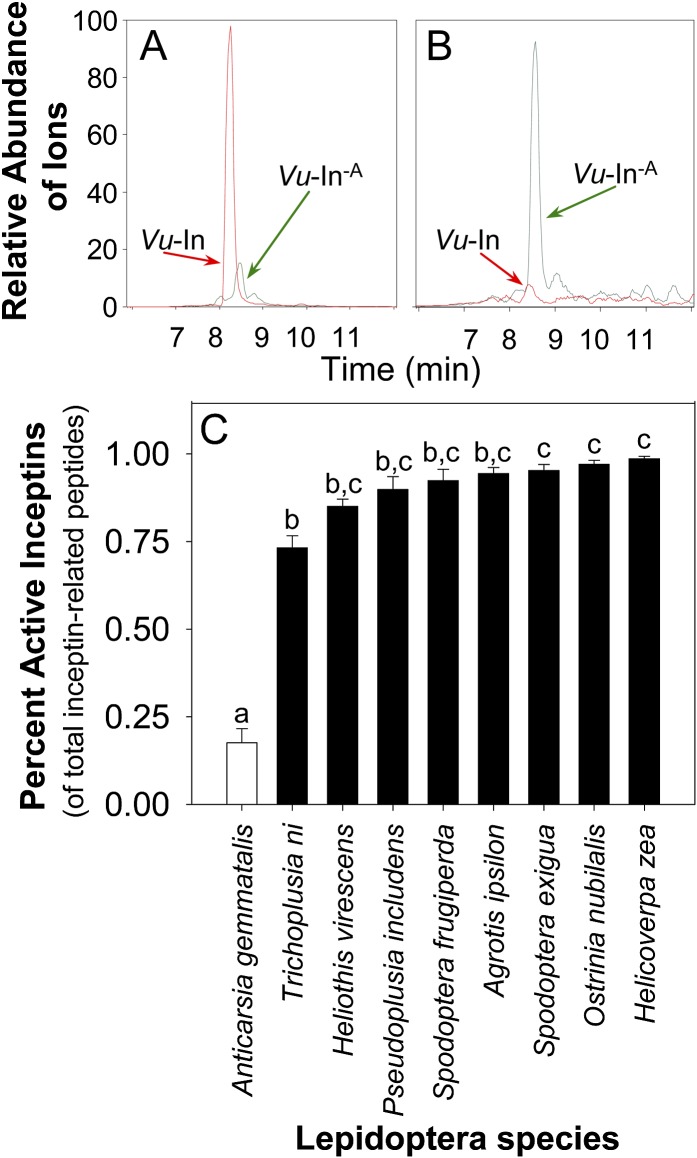
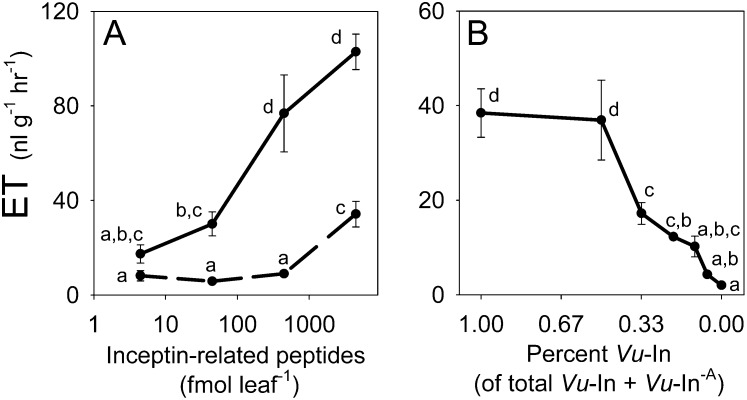
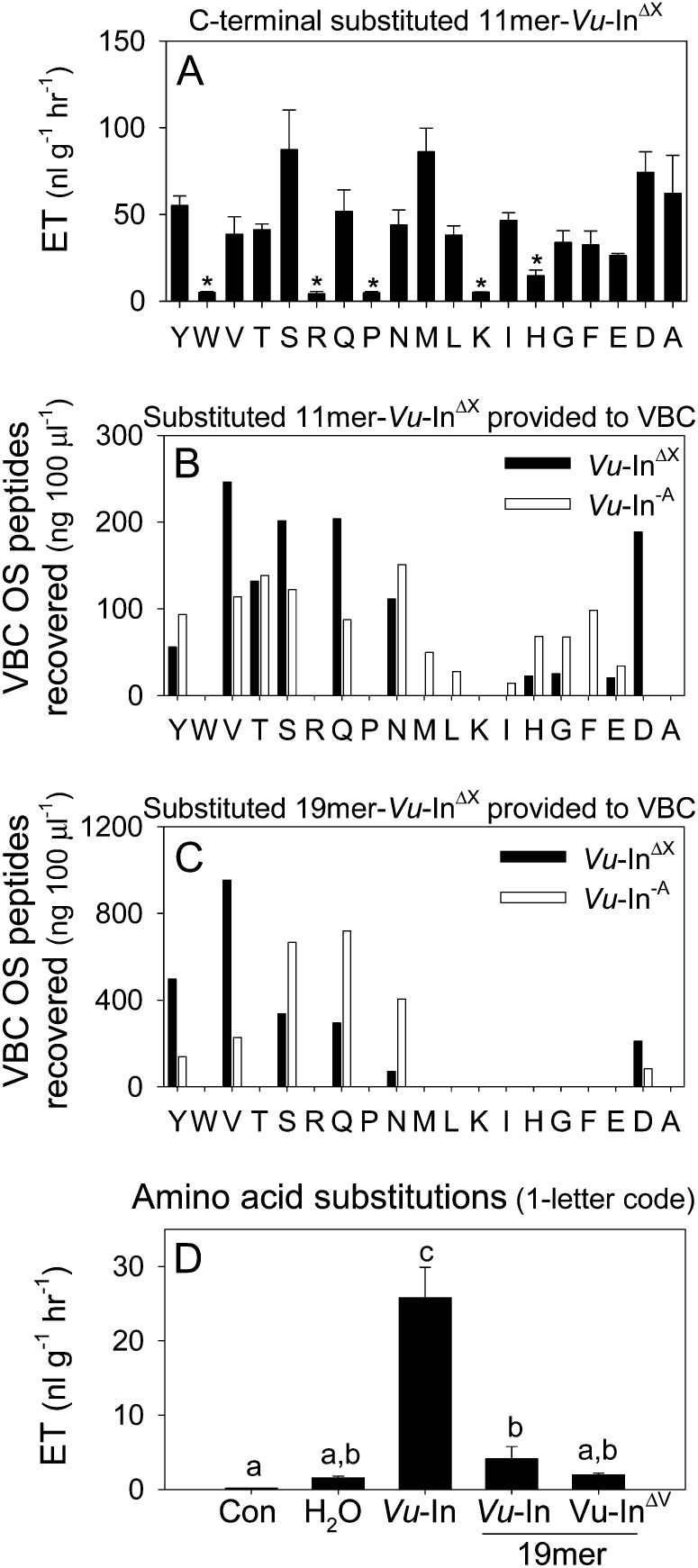
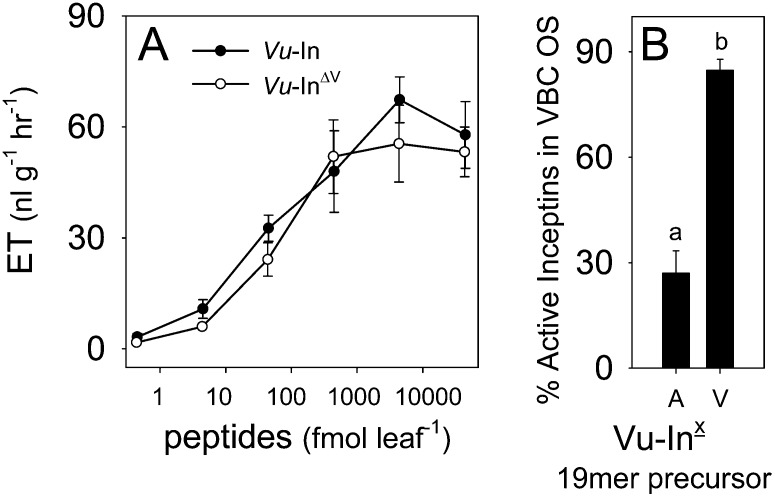
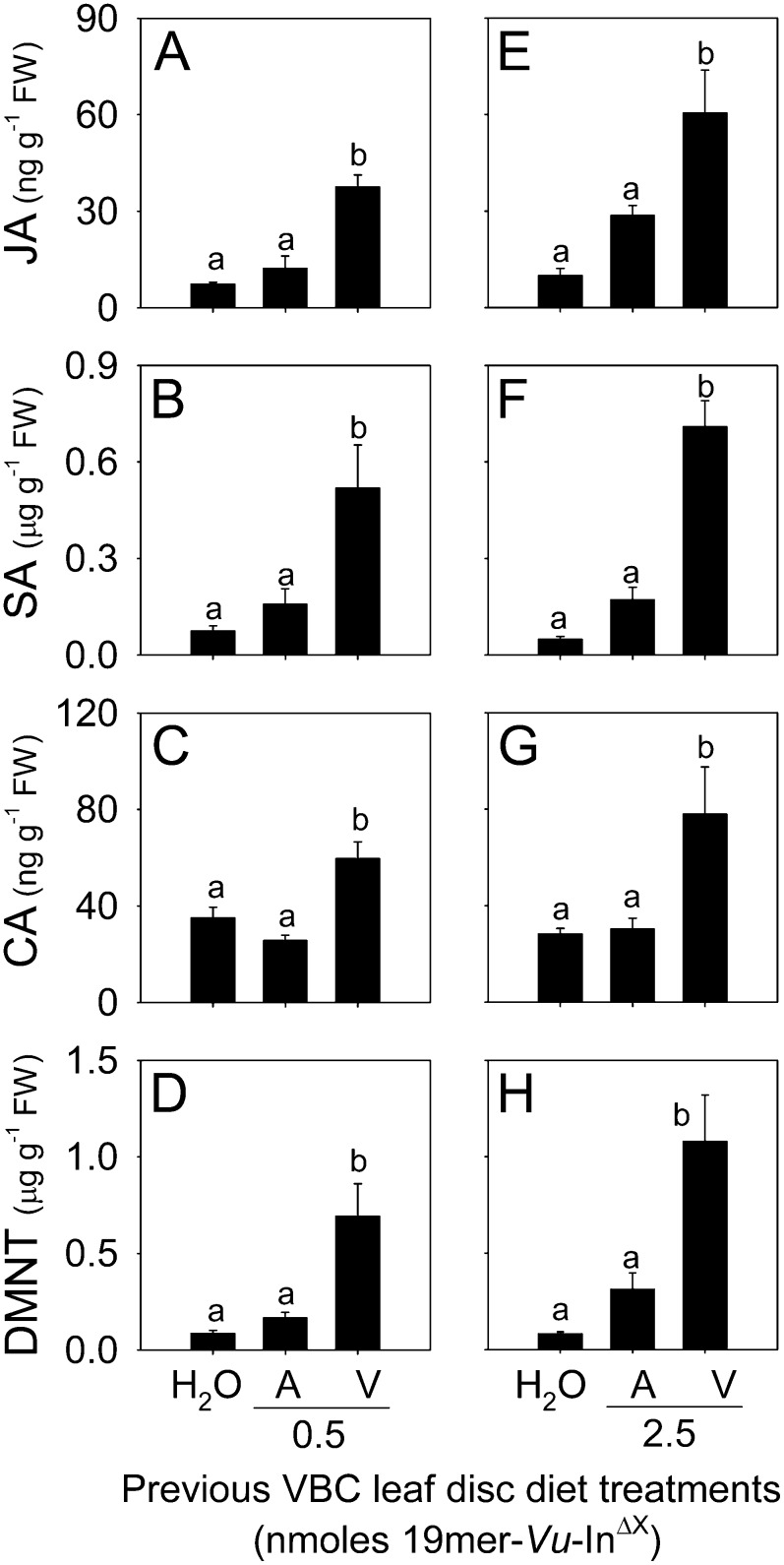
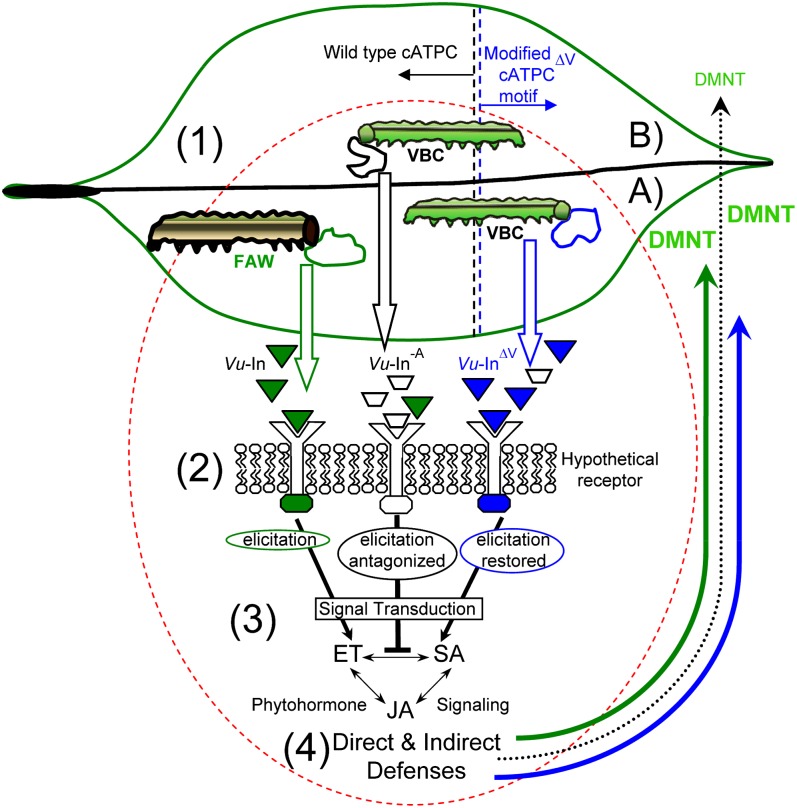
Plants respond to insect herbivory through the production of biochemicals that function as either direct defenses or indirect defenses via the attraction of natural enemies. While attack by closely related insect pests can result in distinctive levels of induced plant defenses, precise biochemical mechanisms responsible for differing responses remain largely unknown. Cowpea (Vigna unguiculata) responds to Fall armyworm (Spodoptera frugiperda) herbivory through the detection of fragments of chloroplastic ATP synthase γ-subunit proteins, termed inceptin-related peptides, present in larval oral secretions (OS). In contrast to generalists like Fall armyworm, OS of the legume-specializing velvetbean caterpillar (VBC; Anticarsia gemmatalis) do not elicit ethylene production and demonstrate significantly lower induced volatile emission in direct herbivory comparisons. Unlike all other Lepidoptera OS examined, which preferentially contain inceptin (Vu-In; +ICDINGVCVDA-), VBC OS contain predominantly a C-terminal truncated peptide, Vu-In(-A) (+ICDINGVCVD-). Vu-In(-A) is both inactive and functions as a potent naturally occurring antagonist of Vu-In-induced responses. To block antagonist production, amino acid substitutions at the C terminus were screened for differences in VBC gut proteolysis. A valine-substituted peptide (Vu-In(ΔV); +ICDINGVCVDV-) retaining full elicitor activity was found to accumulate in VBC OS. Compared with the native polypeptide, VBC that previously ingested 500 pmol of the valine-modified chloroplastic ATP synthase γ-subunit precursor elicited significantly stronger plant responses in herbivory assays. We demonstrate that a specialist herbivore minimizes the activation of defenses by converting an elicitor into an antagonist effector and identify an amino acid substitution that recovers these induced plant defenses to a level observed with generalist herbivores.
Figures
Similar articles
-
Cowpea chloroplastic ATP synthase is the source of multiple plant defense elicitors during insect herbivory.Plant Physiol. 2007 Jun;144(2):793-805. doi: 10.1104/pp.107.097154. Epub 2007 Mar 16. Plant Physiol. 2007. PMID: 17369425 Free PMC article.
-
Fragments of ATP synthase mediate plant perception of insect attack.Proc Natl Acad Sci U S A. 2006 Jun 6;103(23):8894-9. doi: 10.1073/pnas.0602328103. Epub 2006 May 23. Proc Natl Acad Sci U S A. 2006. PMID: 16720701 Free PMC article.
-
The attraction of Spodoptera frugiperda neonates to cowpea seedlings is mediated by volatiles induced by conspecific herbivory and the elicitor inceptin.J Chem Ecol. 2008 Mar;34(3):291-300. doi: 10.1007/s10886-007-9414-y. Epub 2008 Feb 7. J Chem Ecol. 2008. PMID: 18256881
-
Indirect plant defense against insect herbivores: a review.Insect Sci. 2018 Feb;25(1):2-23. doi: 10.1111/1744-7917.12436. Epub 2017 Mar 20. Insect Sci. 2018. PMID: 28035791 Review.
-
The biochemical and molecular mechanisms of plants: a review on insect herbivory.Plant Signal Behav. 2025 Dec;20(1):2439248. doi: 10.1080/15592324.2024.2439248. Epub 2024 Dec 26. Plant Signal Behav. 2025. PMID: 39723993 Review.
Cited by
-
MATI, a Novel Protein Involved in the Regulation of Herbivore-Associated Signaling Pathways.Front Plant Sci. 2017 Jun 9;8:975. doi: 10.3389/fpls.2017.00975. eCollection 2017. Front Plant Sci. 2017. PMID: 28649257 Free PMC article.
-
Planthopper protein Nlsp5 is essential for salivary sheath formation and acts as a HAMP inducing plant resistance to insects.Plant Biotechnol J. 2025 Sep;23(9):4076-4091. doi: 10.1111/pbi.70223. Epub 2025 Jun 23. Plant Biotechnol J. 2025. PMID: 40546084 Free PMC article.
-
Transcriptome profiling revealed novel transcriptional regulators in maize responses to Ostrinia furnacalis and jasmonic acid.PLoS One. 2017 May 16;12(5):e0177739. doi: 10.1371/journal.pone.0177739. eCollection 2017. PLoS One. 2017. PMID: 28520800 Free PMC article.
-
BSR1, a Rice Receptor-like Cytoplasmic Kinase, Positively Regulates Defense Responses to Herbivory.Int J Mol Sci. 2023 Jun 20;24(12):10395. doi: 10.3390/ijms241210395. Int J Mol Sci. 2023. PMID: 37373546 Free PMC article.
-
The Salivary Protein Repertoire of the Polyphagous Spider Mite Tetranychus urticae: A Quest for Effectors.Mol Cell Proteomics. 2016 Dec;15(12):3594-3613. doi: 10.1074/mcp.M116.058081. Epub 2016 Oct 4. Mol Cell Proteomics. 2016. PMID: 27703040 Free PMC article.
References
-
- Adler LS, Schmitt J, Bowers MD. (1995) Genetic variation in defensive chemistry in Plantago lanceolata (Plantaginaceae) and its effect on the specialist herbivore Junonia coenia (Nymphalidae). Oecologia 101: 75–85 - PubMed
-
- Agrawal AA, Kurashige NS. (2003) A role for isothiocyanates in plant resistance against the specialist herbivore Pieris rapae. J Chem Ecol 29: 1403–1415 - PubMed
-
- Alborn HT, Brennan MM, Tumlinson JH. (2003) Differential activity and degradation of plant volatile elicitors in regurgitant of tobacco hornworm (Manduca sexta) larvae. J Chem Ecol 29: 1357–1372 - PubMed
-
- Alborn HT, Turlings TCJ, Jones TH, Stenhagen G, Loughrin JH, Tumlinson JH. (1997) An elicitor of plant volatiles from beet armyworm oral secretion. Science 276: 945–949
-
- Ali JG, Agrawal AA. (2012) Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci 17: 293–302 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
