

Sex differences in microRNA regulation of gene expression: no smoke, just miRs
- PMID: 23009289
- PMCID: PMC3507674
- DOI: 10.1186/2042-6410-3-22
Sex differences in microRNA regulation of gene expression: no smoke, just miRs
Abstract
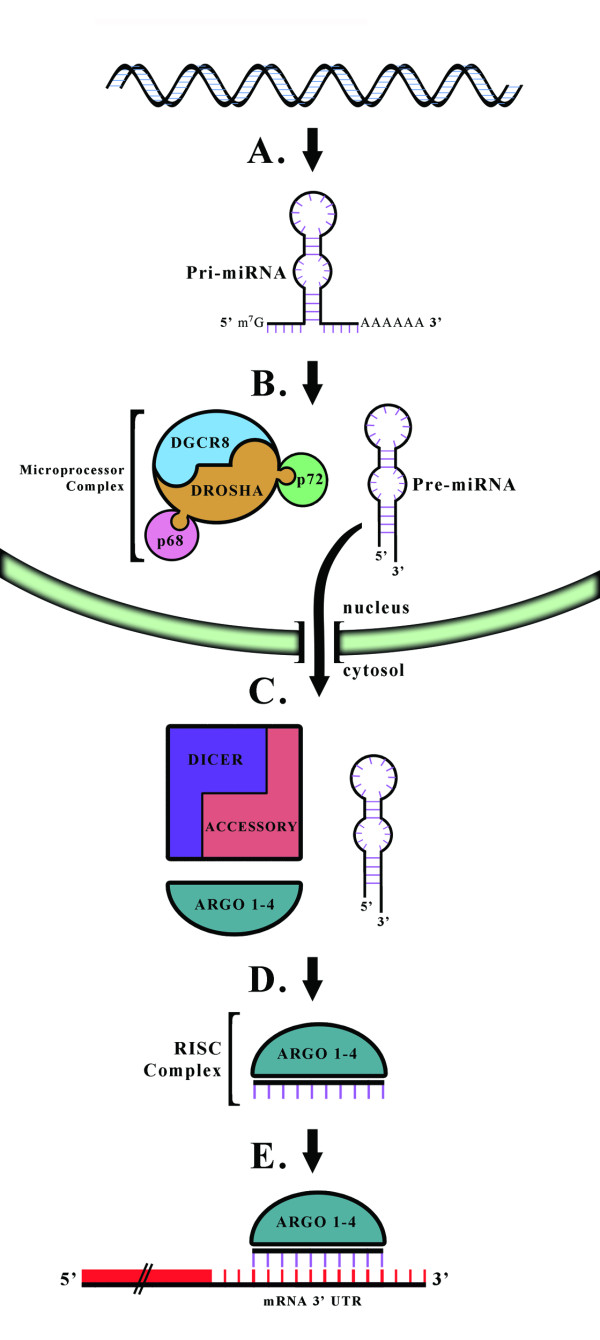
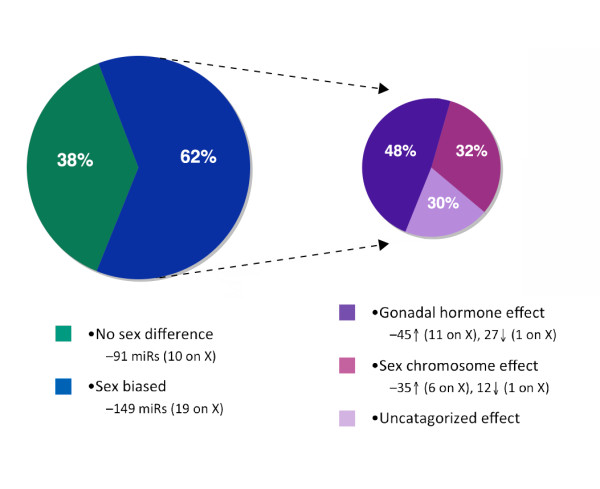
Males and females differ widely in morphology, physiology, and behavior leading to disparities in many health outcomes, including sex biases in the prevalence of many neurodevelopmental disorders. However, with the exception of a relatively small number of genes on the Y chromosome, males and females share a common genome. Therefore, sexual differentiation must in large part be a product of the sex biased expression of this shared genetic substrate. microRNAs (miRs) are small non-coding RNAs involved in the post-transcriptional regulation of up to 70% of protein-coding genes. The ability of miRs to regulate such a vast amount of the genome with a high degree of specificity makes them perfectly poised to play a critical role in programming of the sexually dimorphic brain. This review describes those characteristics of miRs that make them particularly amenable to this task, and examines the influences of both the sex chromosome complement as well as gonadal hormones on their regulation. Exploring miRs in the context of sex differences in disease, particularly in sex-biased neurodevelopmental disorders, may provide novel insight into the pathophysiology and potential therapeutic targets in disease treatment and prevention.
Figures
References
-
- Yeh S-H, Chen P-J. Gender Disparity of Hepatocellular Carcinoma: The Roles of Sex Hormones. Oncology. 2010;78:172–179. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources