

Phosphatidylcholine and the CDP-choline cycle
- PMID: 23010477
- PMCID: PMC3562404
- DOI: 10.1016/j.bbalip.2012.09.009
Phosphatidylcholine and the CDP-choline cycle
Abstract
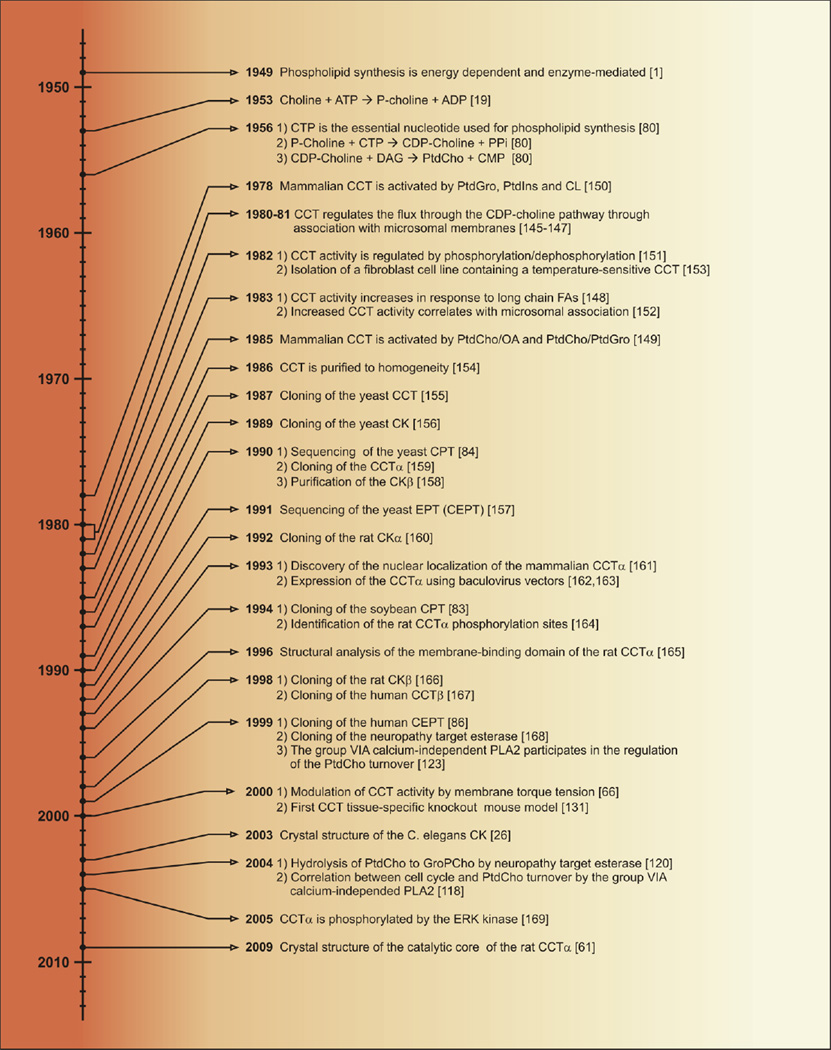
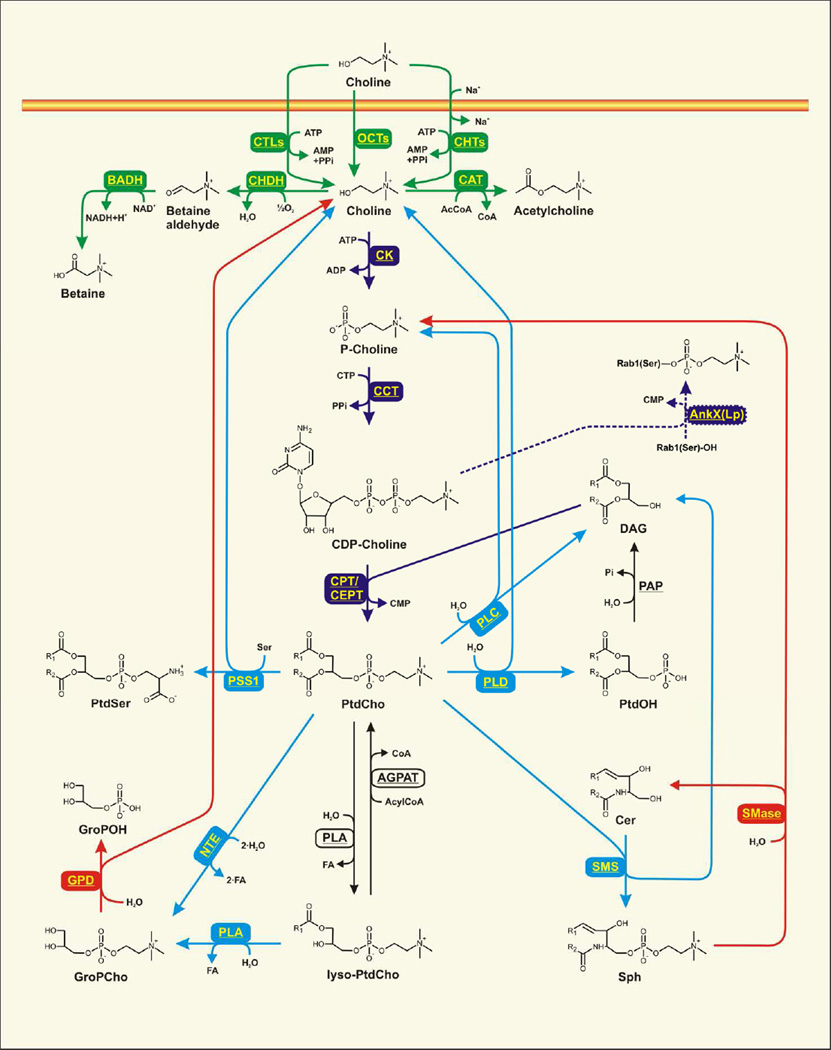
The CDP-choline pathway of phosphatidylcholine (PtdCho) biosynthesis was first described more than 50 years ago. Investigation of the CDP-choline pathway in yeast provides a basis for understanding the CDP-choline pathway in mammals. PtdCho is considered as an intermediate in a cycle of synthesis and degradation, and the activity of a CDP-choline cycle is linked to subcellular membrane lipid movement. The components of the mammalian CDP-choline pathway include choline transport, choline kinase, phosphocholine cytidylyltransferase, and choline phosphotransferase activities. The protein isoforms and biochemical mechanisms of regulation of the pathway enzymes are related to their cell- and tissue-specific functions. Regulated PtdCho turnover mediated by phospholipases or neuropathy target esterase participates in the mammalian CDP-choline cycle. Knockout mouse models define the biological functions of the CDP-choline cycle in mammalian cells and tissues. This article is part of a Special Issue entitled Phospholipids and Phospholipid Metabolism.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Friedkin M, Lehninger AL. Oxidation-coupled incorporation of inorganic radiophosphate into phospholipide and nucleic acid in a cell-free system. J. Biol. Chem. 1949;177:775–788. - PubMed
-
- Kennedy EP. Metabolism of lipides. Annu. Rev. Biochem. 1957;26:119–148. - PubMed
-
- Vance JE, Vance DE. Phospholipid biosynthesis in mammalian cells. Biochem. Cell Biol. 2004;82:113–128. - PubMed
-
- Gibellini F, Smith TK. The Kennedy pathway-De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB. Life. 2010;62:414–428. - PubMed
-
- Stalberg K, Neal AC, Ronne H, Stahl U. Identification of a novel GPCAT activity and a new pathway for phosphatidylcholine biosynthesis in S. cerevisiae. J. Lipid Res. 2008;49:1794–1806. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
