

Hyaluronan activation of the Nlrp3 inflammasome contributes to the development of airway hyperresponsiveness
- PMID: 23010656
- PMCID: PMC3546367
- DOI: 10.1289/ehp.1205188
Hyaluronan activation of the Nlrp3 inflammasome contributes to the development of airway hyperresponsiveness
Retraction in
-
Retraction: Hyaluronan activation of the Nlrp3 inflammasome contributes to the development of airway hyperresponsiveness.Environ Health Perspect. 2015 Jul;123(7):A172. doi: 10.1289/ehp.1205188RET. Environ Health Perspect. 2015. PMID: 26133028 Free PMC article. No abstract available.
Abstract
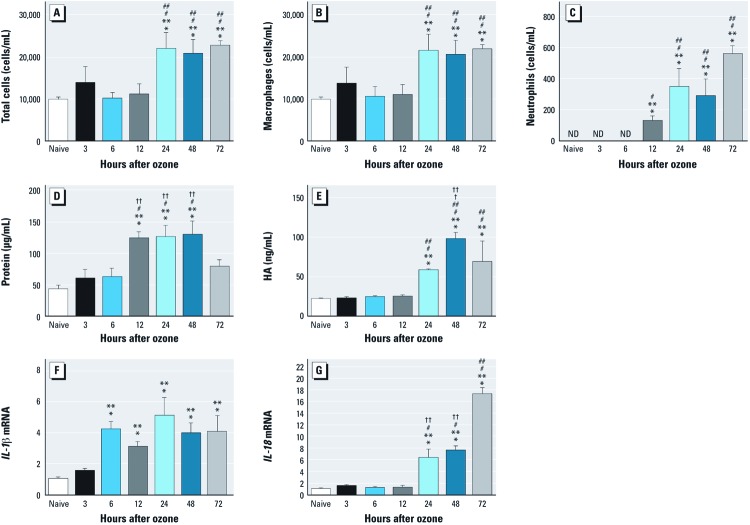
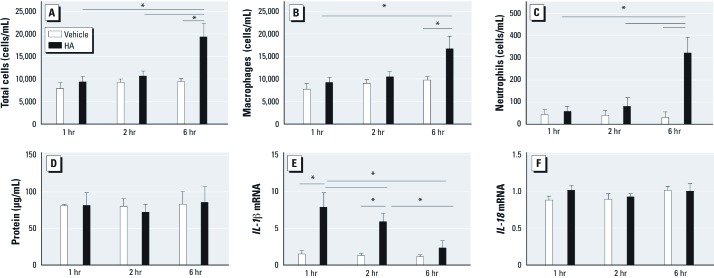
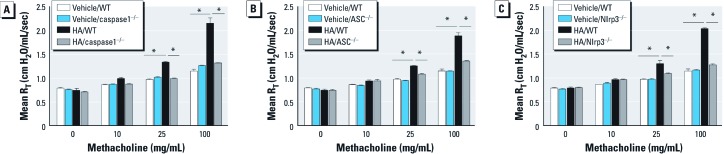
Background: The role of the Nlrp3 inflammasome in nonallergic airway hyperresponsiveness (AHR) has not previously been reported. Recent evidence supports both interleukin (IL) 1β and short fragments of hyaluronan (HA) as contributors to the biological response to inhaled ozone.
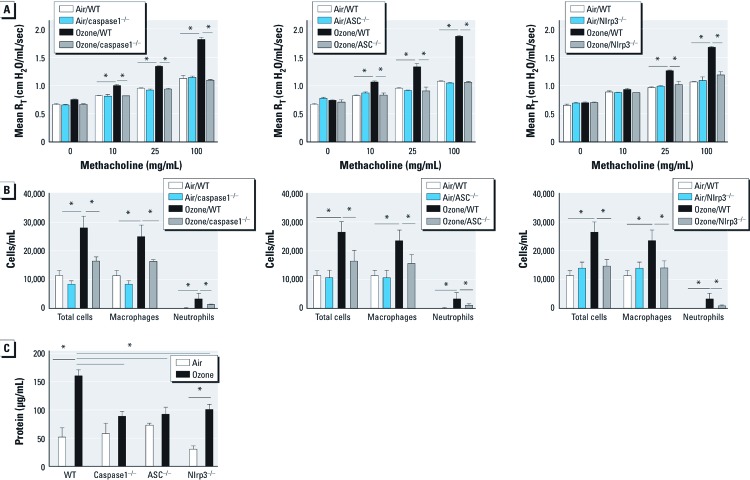
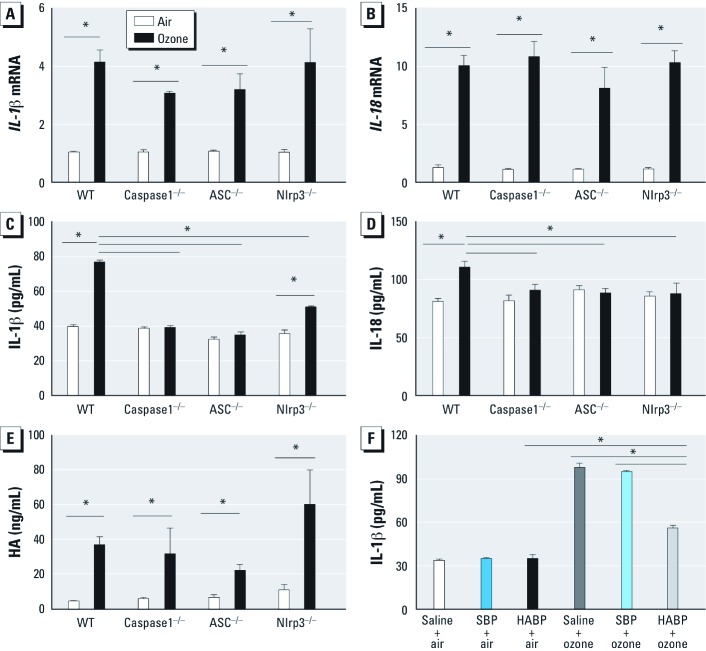
Objective: Because extracellular secretion of IL-1β requires activation of the inflammasome, we investigated the role of the inflammasome proteins ASC, caspase1, and Nlrp3 in the biological response to ozone and HA.
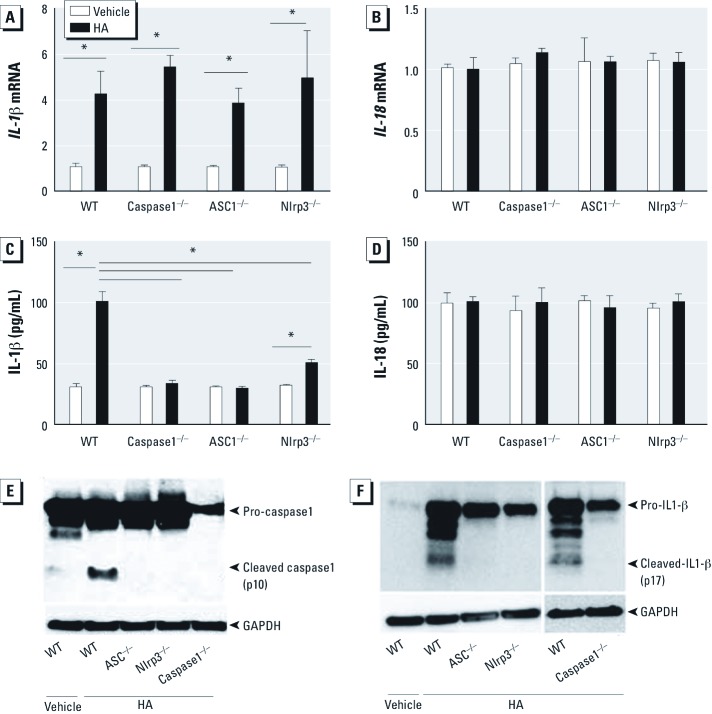
Methods: C57BL/6J wild-type mice and mice deficient in ASC, caspase1, or Nlrp3 were exposed to ozone (1 ppm for 3 hr) or HA followed by analysis of airway resistance, cellular inflammation, and total protein and cytokines in bronchoalveolar lavage fluid (BALF). Transcription levels of IL-1β and IL-18 were determined in two populations of lung macrophages. In addition, we examined levels of cleaved caspase1 and cleaved IL-1β as markers of inflammasome activation in isolated alveolar macrophages harvested from BALF from HA-treated mice.
Results: We observed that genes of the Nlrp3 inflammasome were required for development of AHR following exposure to either ozone or HA fragments. These genes are partially required for the cellular inflammatory response to ozone. The expression of IL-1β mRNA in alveolar macrophages was up-regulated after either ozone or HA challenge and was not dependent on the Nlrp3 inflammasome. However, soluble levels of IL-1β protein were dependent on the inflammasome after challenge with either ozone or HA. HA challenge resulted in cleavage of macrophage-derived caspase1 and IL-1β, suggesting a role for alveolar macrophages in Nlrp3-dependent AHR.
Conclusions: The Nlrp3 inflammasome is required for the development of ozone-induced reactive airways disease.
Conflict of interest statement
The authors declare they have no actual or potential competing financial interests.
Figures
Comment in
-
Findings of Research Misconduct.Fed Regist. 2019 Nov 7;84(216):60097-60098. Fed Regist. 2019. PMID: 37547121 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous