

LacZ β-galactosidase: structure and function of an enzyme of historical and molecular biological importance
- PMID: 23011886
- PMCID: PMC3575911
- DOI: 10.1002/pro.2165
LacZ β-galactosidase: structure and function of an enzyme of historical and molecular biological importance
Abstract
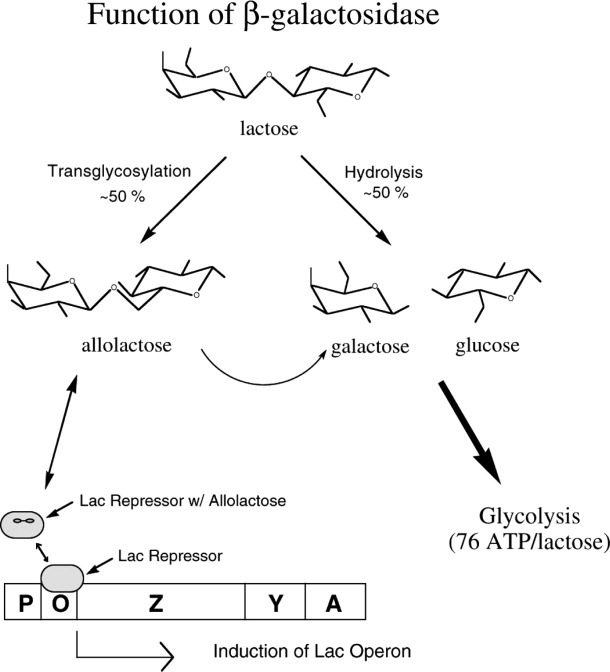
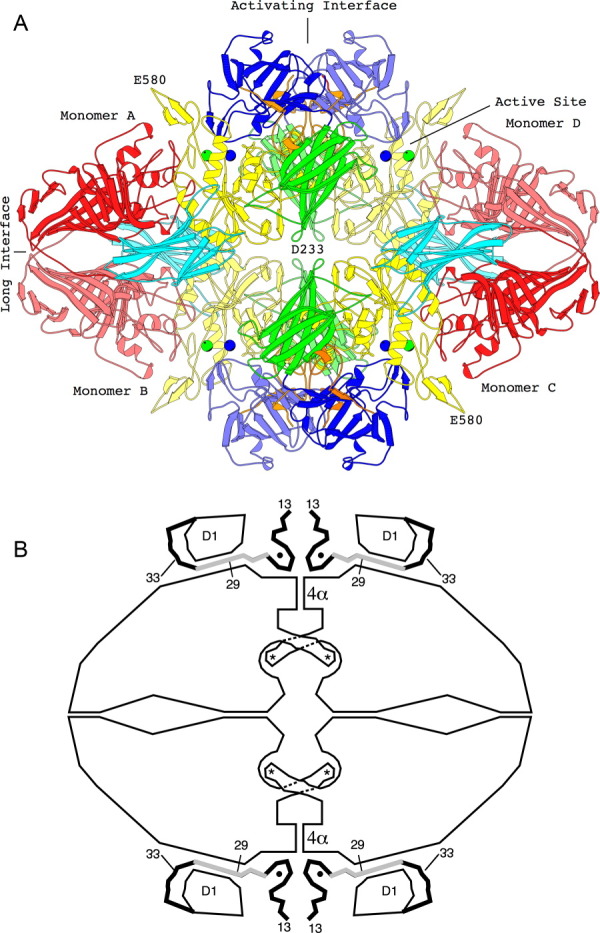
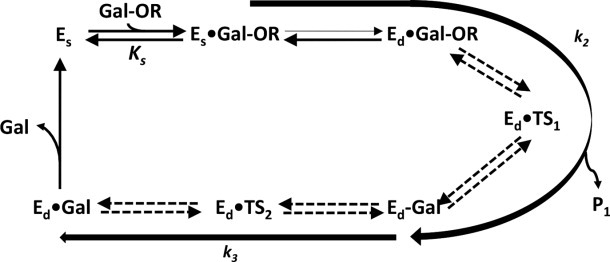
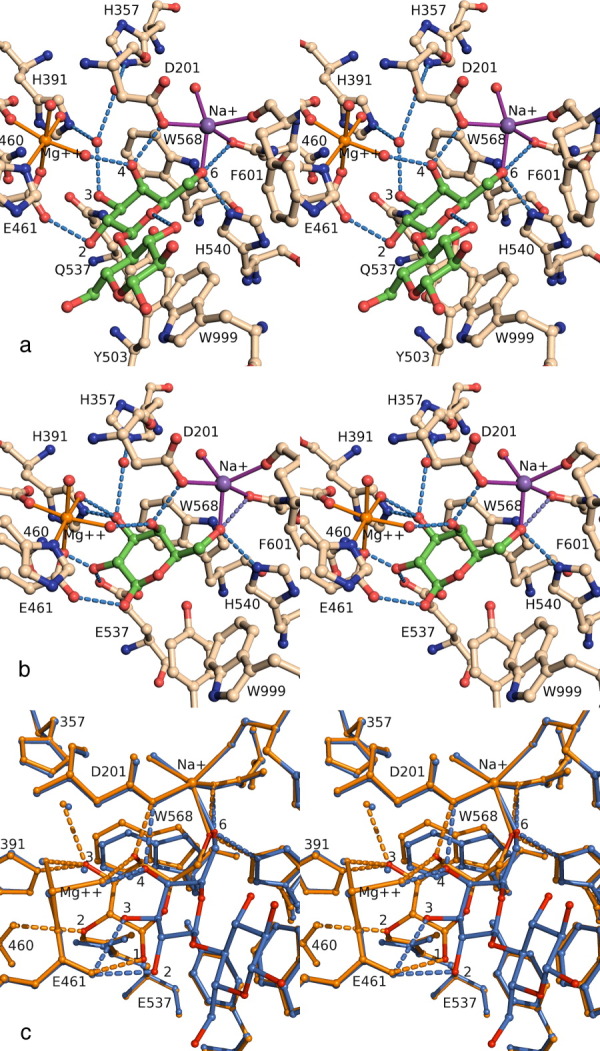
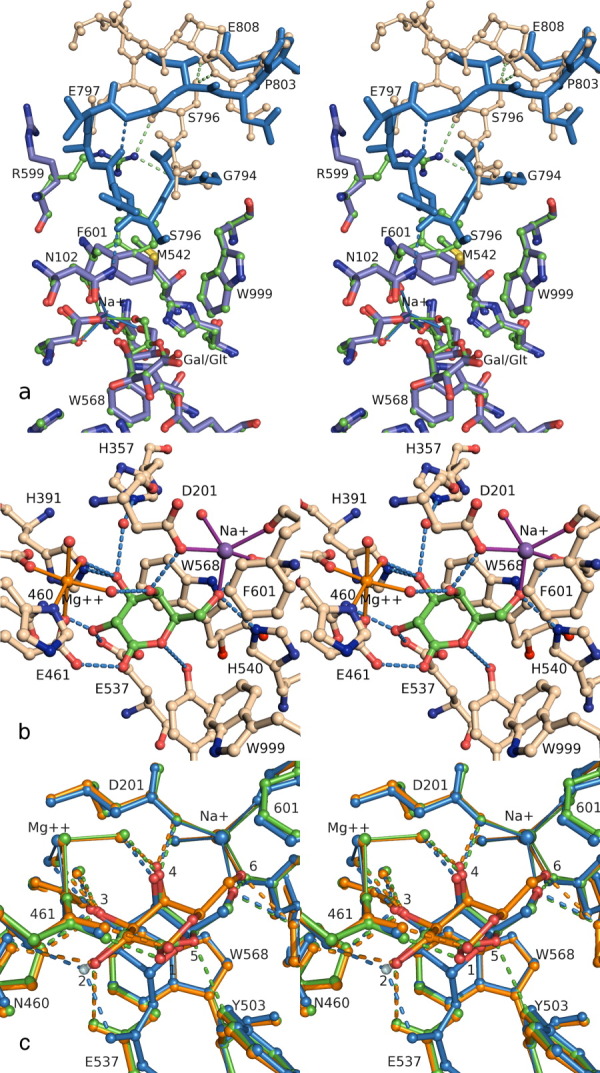
This review provides an overview of the structure, function, and catalytic mechanism of lacZ β-galactosidase. The protein played a central role in Jacob and Monod's development of the operon model for the regulation of gene expression. Determination of the crystal structure made it possible to understand why deletion of certain residues toward the amino-terminus not only caused the full enzyme tetramer to dissociate into dimers but also abolished activity. It was also possible to rationalize α-complementation, in which addition to the inactive dimers of peptides containing the "missing" N-terminal residues restored catalytic activity. The enzyme is well known to signal its presence by hydrolyzing X-gal to produce a blue product. That this reaction takes place in crystals of the protein confirms that the X-ray structure represents an active conformation. Individual tetramers of β-galactosidase have been measured to catalyze 38,500 ± 900 reactions per minute. Extensive kinetic, biochemical, mutagenic, and crystallographic analyses have made it possible to develop a presumed mechanism of action. Substrate initially binds near the top of the active site but then moves deeper for reaction. The first catalytic step (called galactosylation) is a nucleophilic displacement by Glu537 to form a covalent bond with galactose. This is initiated by proton donation by Glu461. The second displacement (degalactosylation) by water or an acceptor is initiated by proton abstraction by Glu461. Both of these displacements occur via planar oxocarbenium ion-like transition states. The acceptor reaction with glucose is important for the formation of allolactose, the natural inducer of the lac operon.
Copyright © 2012 The Protein Society.
Figures
References
-
- Jacob F, Monod J. Genetic regulatory mechanisms in the synthesis of proteins. J Mol Biol. 1961;3:318–356. - PubMed
-
- Huber RE, Kurz G, Wallenfels K. A quantitation of the factors which affect the hyrolase and trangalactosylase activities of β-galactosidase (E. coli) on lactose. Biochemistry. 1976;15:1994–2001. - PubMed
-
- Wyckoff HW, Doscher M, Tsernoglou D, Inagami T, Johnson LN, Hardman KD, Allewell NM, Kelly DM, Richards FM. Design of a diffractometer and flow cell system for X-ray analysis of crystalline proteins with applications to the crystal chemistry of ribonuclease-S. J Mol Biol. 1967;27:563–578. - PubMed
-
- Matthews BW. Determination of molecular weight from protein crystals. J Mol Biol. 1974;82:513–526. - PubMed
-
- Fowler A, Zabin I. Amino acid sequence of β-galactosidase. XI. Peptide ordering procedures and the complete sequence. J Biol Chem. 1978;253:5521–5525. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
