

Interactions of the CpxA sensor kinase and cognate CpxR response regulator from Yersinia pseudotuberculosis
- PMID: 23013530
- PMCID: PMC3517363
- DOI: 10.1186/1756-0500-5-536
Interactions of the CpxA sensor kinase and cognate CpxR response regulator from Yersinia pseudotuberculosis
Abstract
Background: The CpxA sensor kinase-CpxR response regulator two-component regulatory system is a sentinel of bacterial envelope integrity. Integrating diverse signals, it can alter the expression of a wide array of components that serve to shield the envelope from damage and to promote bacterial survival. In bacterial pathogens such as Yersinia pseudotuberculosis, this also extends to pathogenesis. CpxR is thought to dimerize upon phosphorylation by the sensor kinase CpxA. This phosphorylation enables CpxR binding to specific DNA sequences where it acts on gene transcription. As Cpx pathway activation is dependent on protein-protein interactions, we performed an interaction analysis of CpxR and CpxA from Y. pseudotuberculosis.
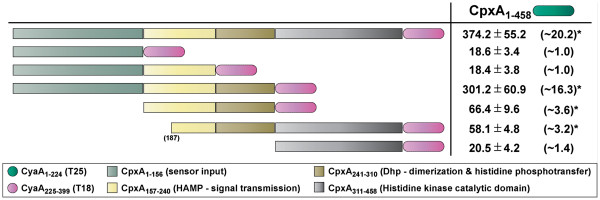
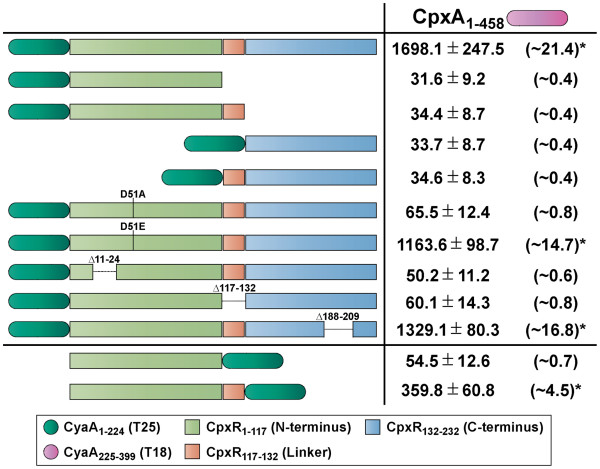
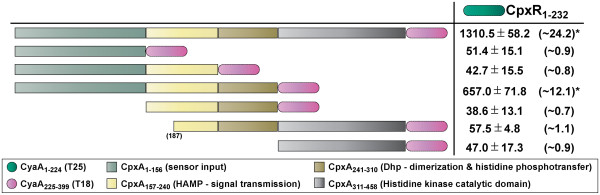
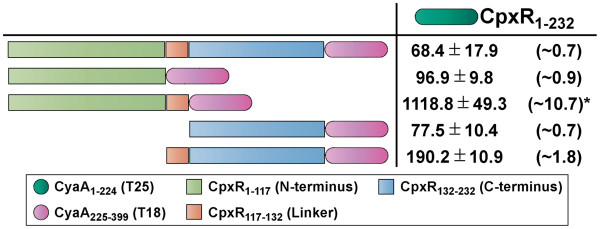
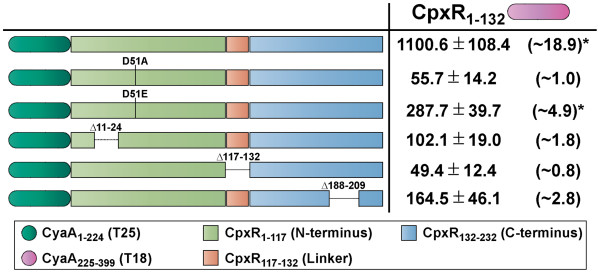
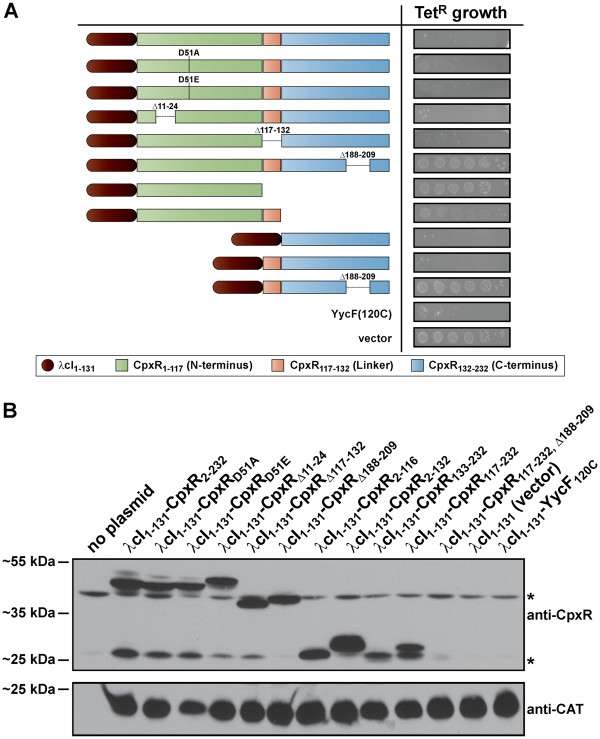
Results: CpxR full-length and truncated versions that either contained or lacked a putative internal linker were all assessed for their ability to homodimerize and interact with CpxA. Using an adenylate cyclase-based bacterial two hybrid approach, full-length CpxR readily engaged with CpxA. The CpxR N-terminus could also homodimerize with itself and with a full-length CpxR. A second homodimerization assay based upon the λcI repressor also demonstrated that the CpxR C-terminus could homodimerize. While the linker was not specifically required, it enhanced CpxR homodimerization. Mutagenesis of cpxR identified the aspartate at residue 51, putative N-terminal coiled-coil and C-terminal winged-helix-turn-helix domains as mediators of CpxR homodimerization. Scrutiny of CpxA full-length and truncated versions revealed that dimerization involved the N-terminus and an internal dimerization and histidine phosphotransfer domain.
Conclusions: This interaction analysis mapped regions of CpxR and CpxA that were responsible for interactions with self or with each other. When combined with other physiological and biochemical tests both hybrid-based assays can be useful in dissecting molecular contacts that may underpin Cpx pathway activation and repression.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
