

Activation of autoreactive B cells by endogenous TLR7 and TLR3 RNA ligands
- PMID: 23019335
- PMCID: PMC3501070
- DOI: 10.1074/jbc.M112.383000
Activation of autoreactive B cells by endogenous TLR7 and TLR3 RNA ligands
Abstract
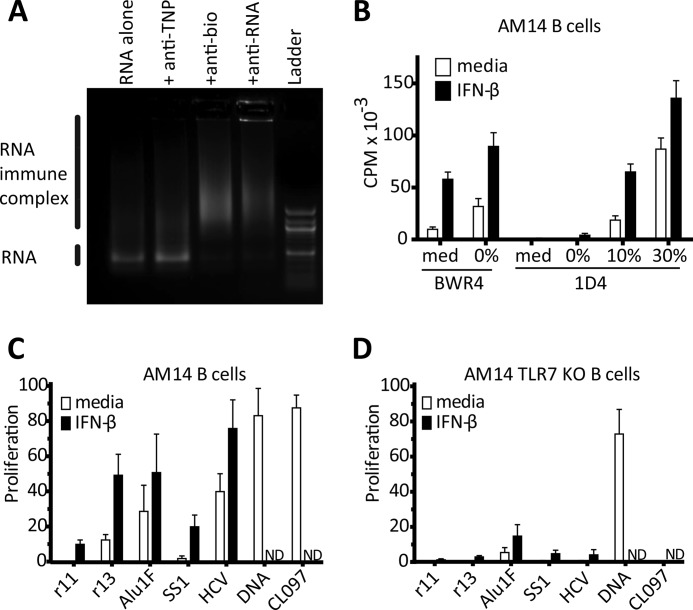
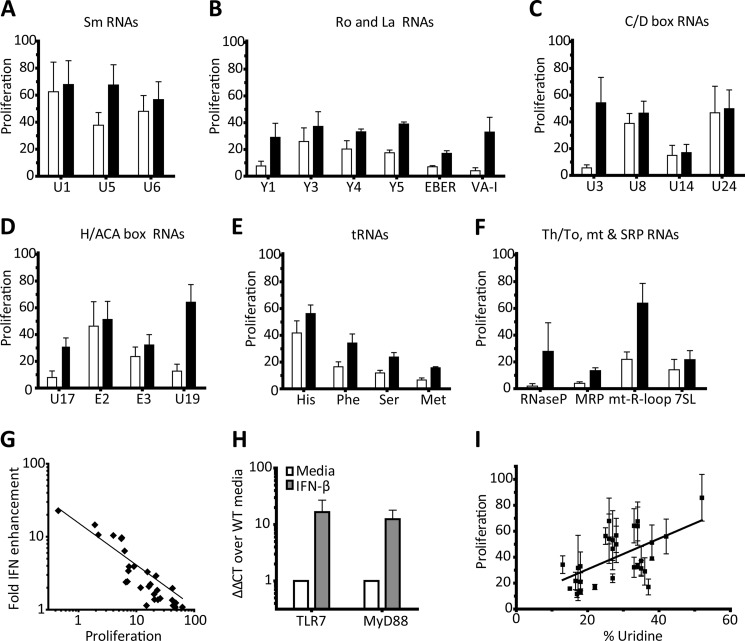
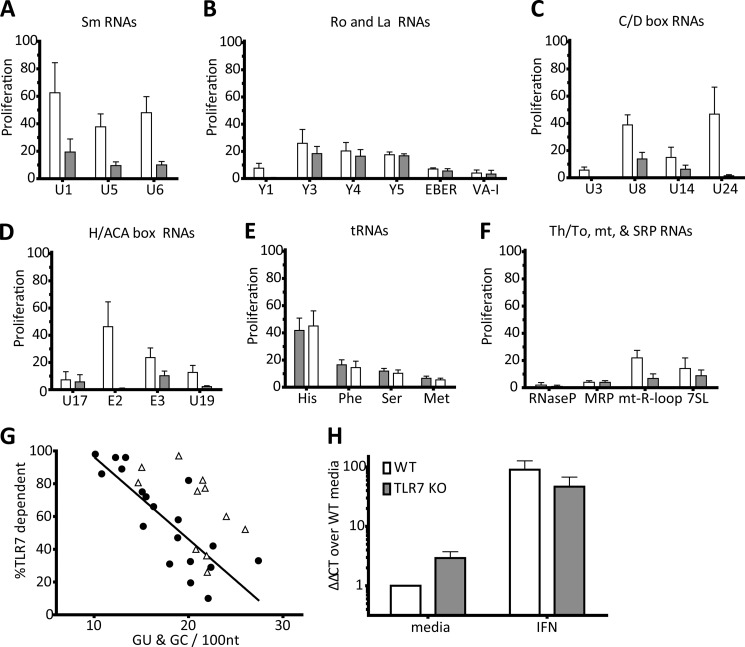
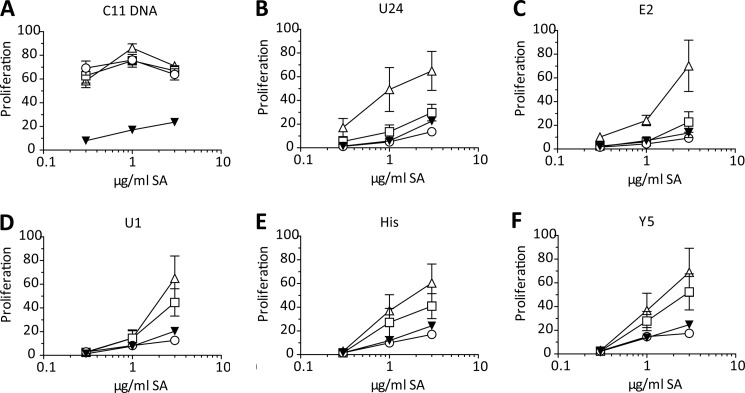
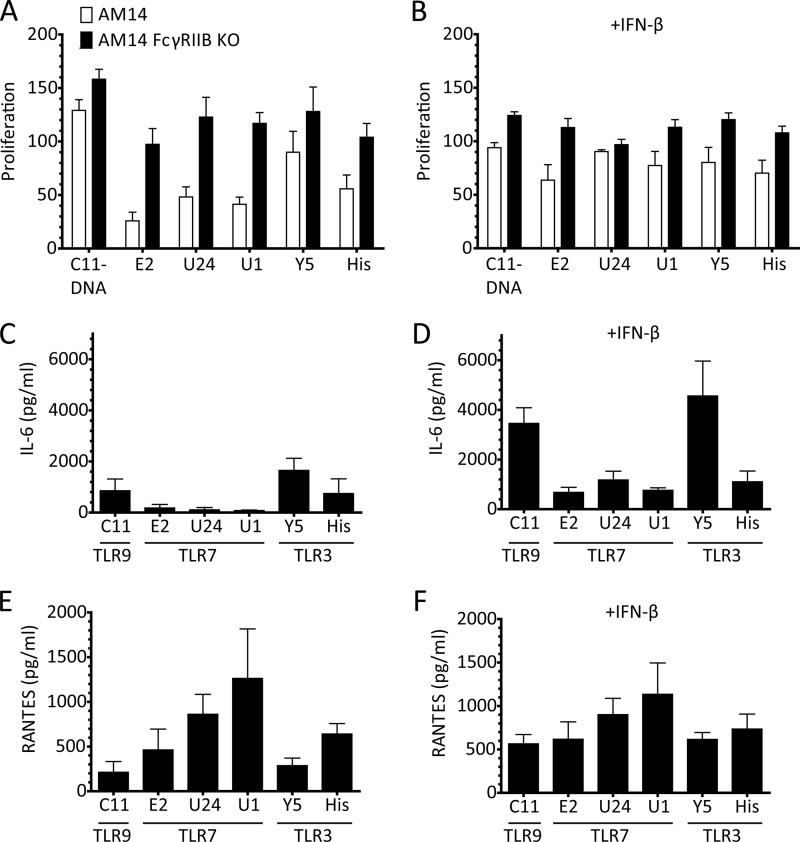
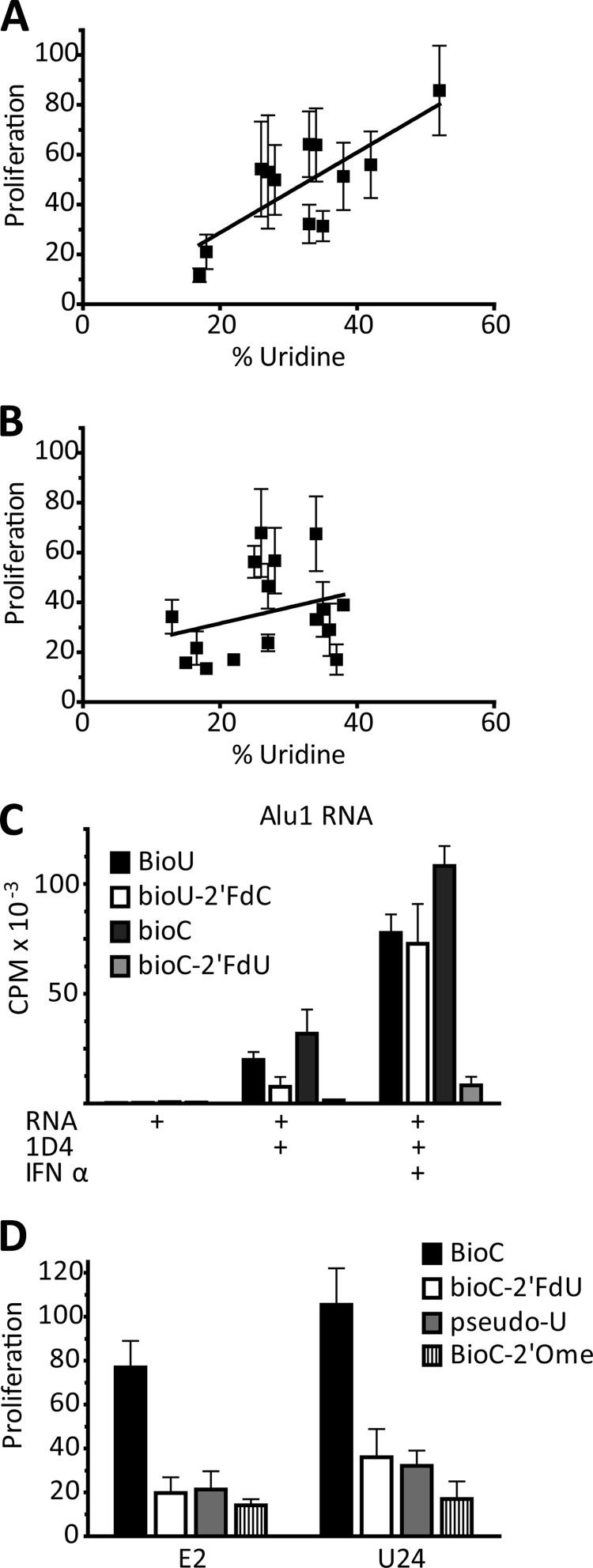
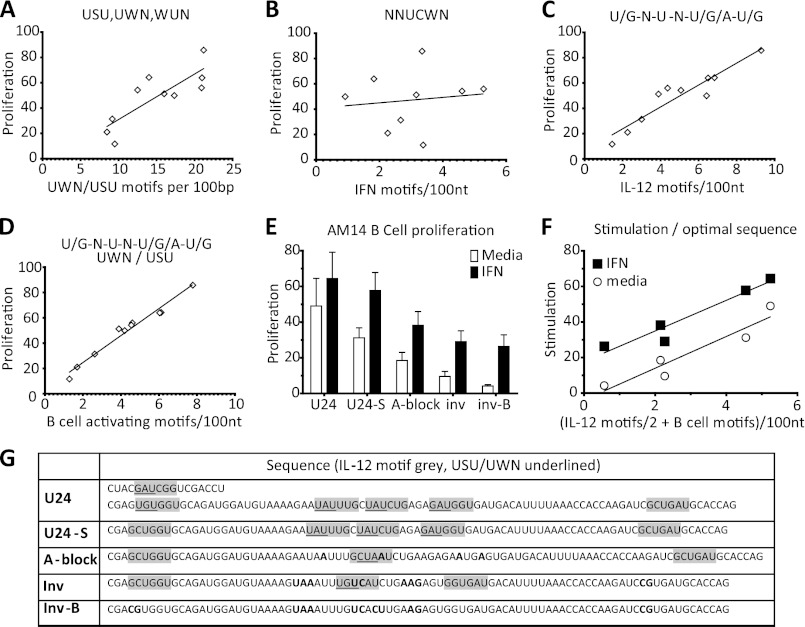
The key step in the activation of autoreactive B cells is the internalization of nucleic acid containing ligands and delivery of these ligands to the Toll-like Receptor (TLR) containing endolysosomal compartment. Ribonucleoproteins represent a large fraction of autoantigens in systemic autoimmune diseases. Here we demonstrate that many uridine-rich mammalian RNA sequences associated with common autoantigens effectively activate autoreactive B cells. Priming with type I IFN increased the magnitude of activation, and the range of which RNAs were stimulatory. A subset of RNAs that contain a high degree of self-complementarity also activated B cells through TLR3. For the RNA sequences that activated predominantly through TLR7, the activation is proportional to uridine-content, and more precisely defined by the frequency of specific uridine-containing motifs. These results identify parameters that define specific mammalian RNAs as ligands for TLRs.
Figures
References
-
- Tan E. M., Cohen A. S., Fries J. F., Masi A. T., McShane D. J., Rothfield N. F., Schaller J. G., Talal N., Winchester R. J. (1982) The 1982 revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 25, 1271–1277 - PubMed
-
- Harris M. L., Rosen A. (2003) Autoimmunity in scleroderma: the origin, pathogenetic role, and clinical significance of autoantibodies. Current Opin. Rheumatol. 15, 778–784 - PubMed
-
- Levine S. M., Raben N., Xie D., Askin F. B., Tuder R., Mullins M., Rosen A., Casciola-Rosen L. A. (2007) Novel conformation of histidyl-transfer RNA synthetase in the lung: the target tissue in Jo-1 autoantibody-associated myositis. Arthritis Rheum. 56, 2729–2739 - PubMed
-
- Lau C. M., Broughton C., Tabor A. S., Akira S., Flavell R. A., Mamula M. J., Christensen S. R., Shlomchik M. J., Viglianti G. A., Rifkin I. R., Marshak-Rothstein A. (2005) RNA-associated autoantigens activate B cells by combined B cell antigen receptor/Toll-like receptor 7 engagement. J. Exp. Med. 202, 1171–1177 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
