

Evidence that viral RNAs have evolved for efficient, two-stage packaging
- PMID: 23019360
- PMCID: PMC3465389
- DOI: 10.1073/pnas.1204357109
Evidence that viral RNAs have evolved for efficient, two-stage packaging
Abstract
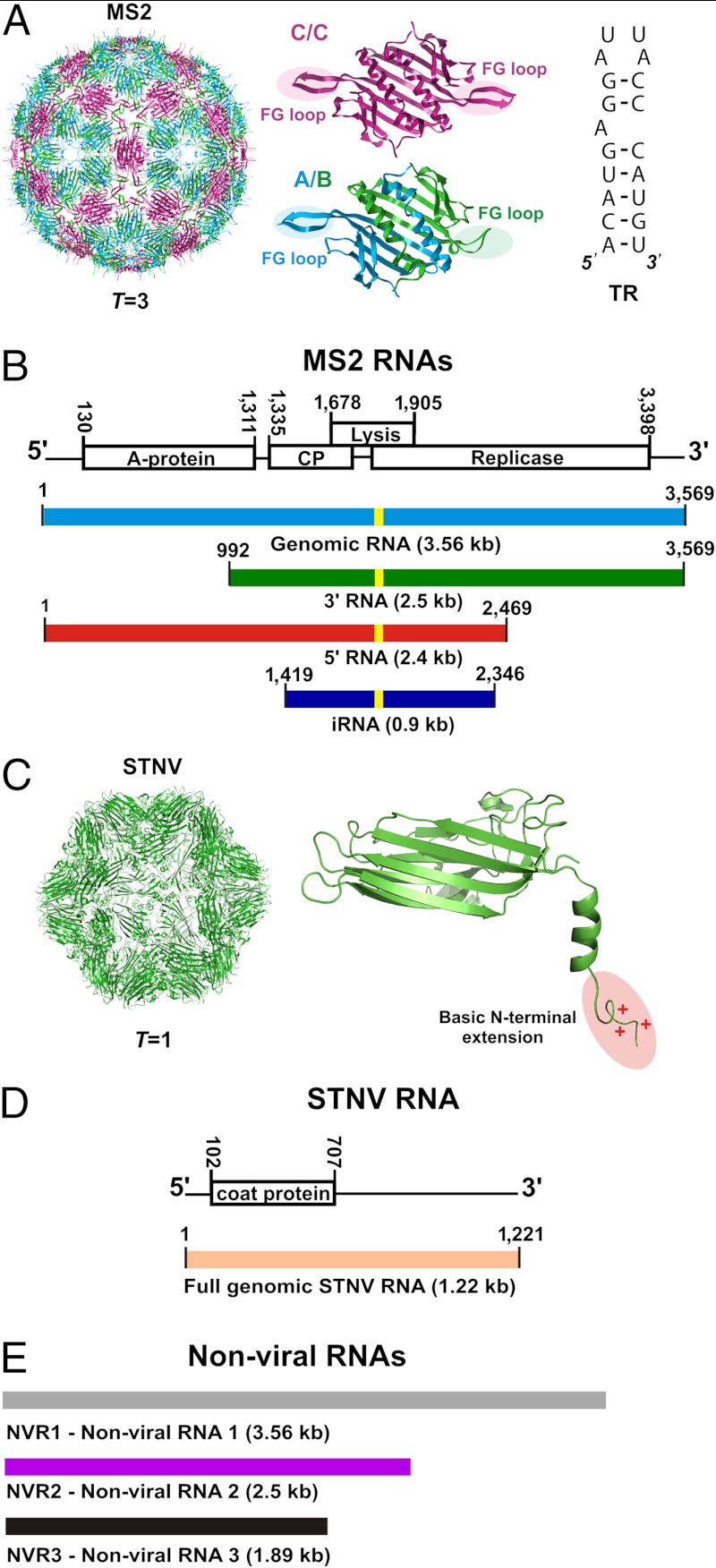
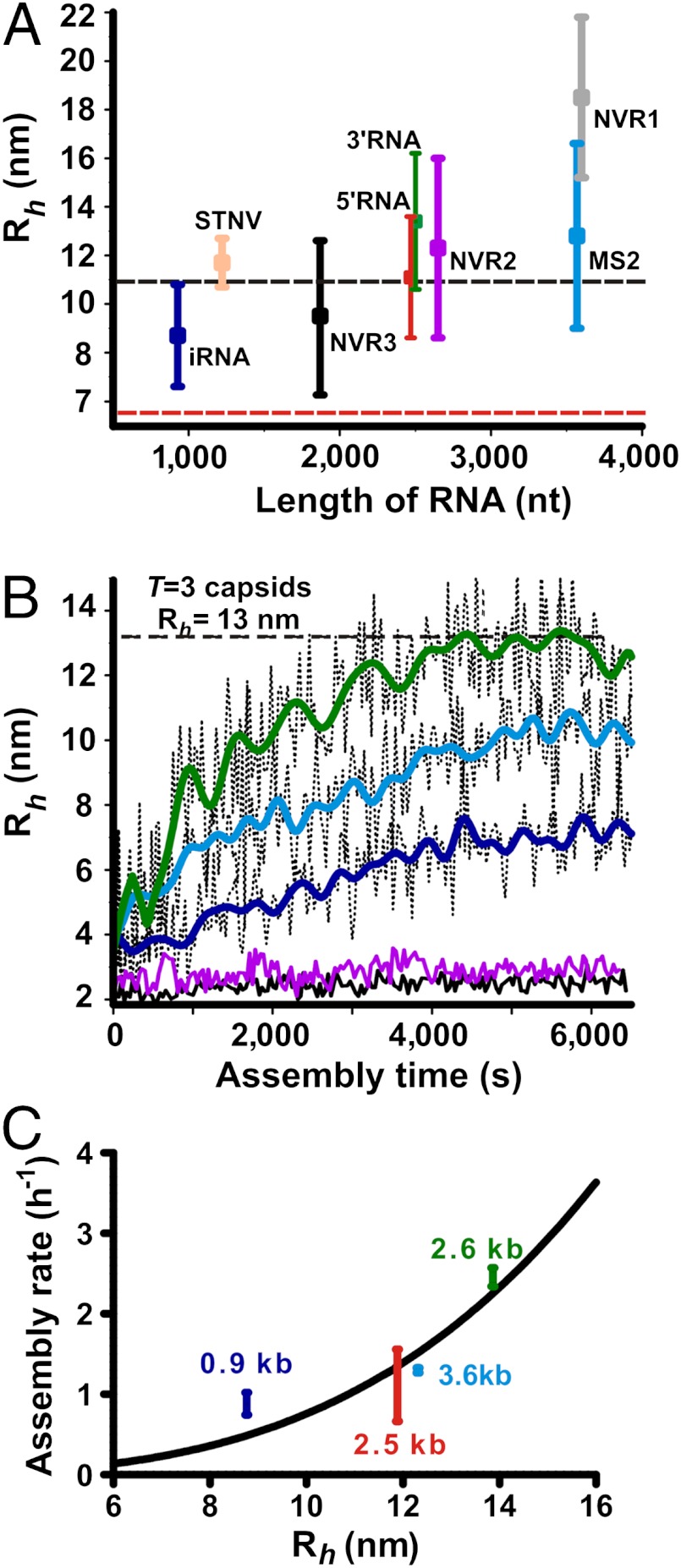
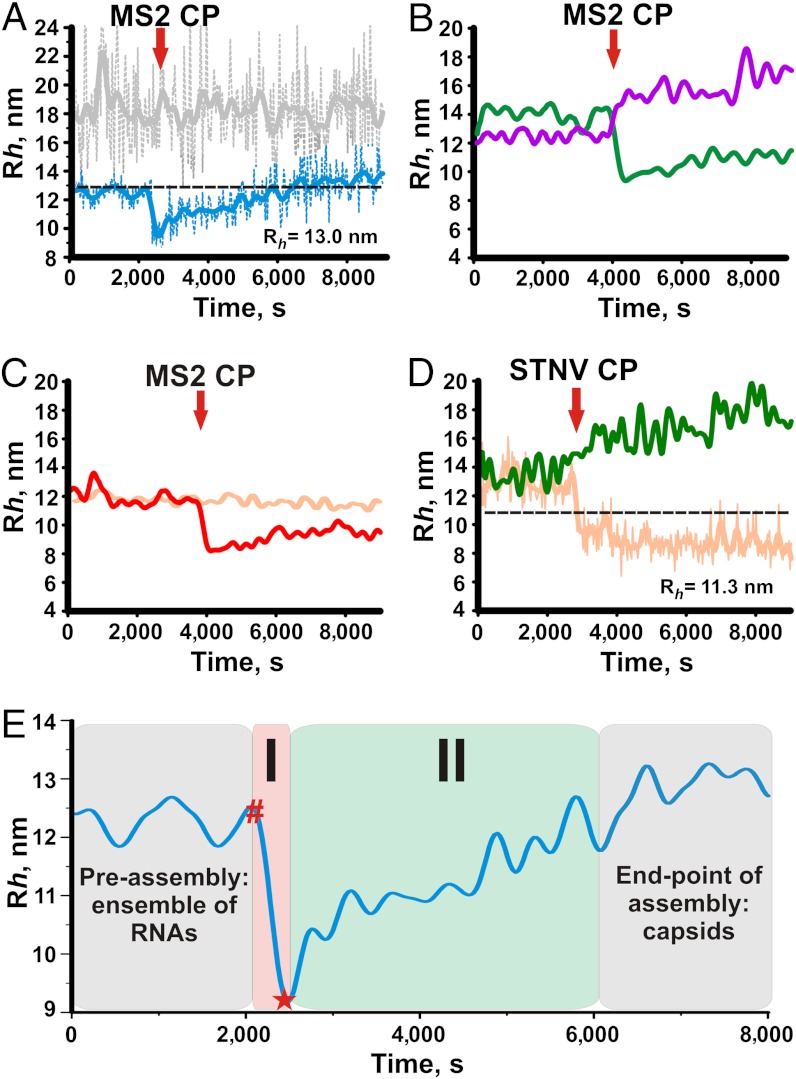
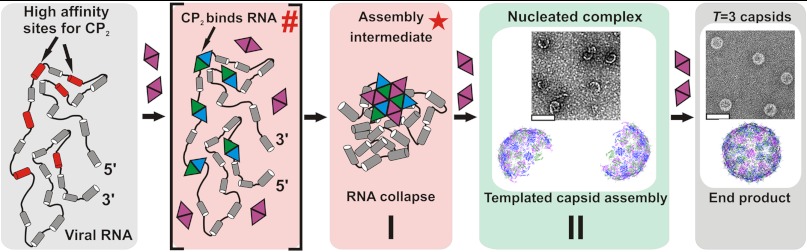
Genome packaging is an essential step in virus replication and a potential drug target. Single-stranded RNA viruses have been thought to encapsidate their genomes by gradual co-assembly with capsid subunits. In contrast, using a single molecule fluorescence assay to monitor RNA conformation and virus assembly in real time, with two viruses from differing structural families, we have discovered that packaging is a two-stage process. Initially, the genomic RNAs undergo rapid and dramatic (approximately 20-30%) collapse of their solution conformations upon addition of cognate coat proteins. The collapse occurs with a substoichiometric ratio of coat protein subunits and is followed by a gradual increase in particle size, consistent with the recruitment of additional subunits to complete a growing capsid. Equivalently sized nonviral RNAs, including high copy potential in vivo competitor mRNAs, do not collapse. They do support particle assembly, however, but yield many aberrant structures in contrast to viral RNAs that make only capsids of the correct size. The collapse is specific to viral RNA fragments, implying that it depends on a series of specific RNA-protein interactions. For bacteriophage MS2, we have shown that collapse is driven by subsequent protein-protein interactions, consistent with the RNA-protein contacts occurring in defined spatial locations. Conformational collapse appears to be a distinct feature of viral RNA that has evolved to facilitate assembly. Aspects of this process mimic those seen in ribosome assembly.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
