

Antioxidant mimetics modulate oxidative stress and cellular signaling in airway epithelial cells infected with respiratory syncytial virus
- PMID: 23023968
- PMCID: PMC3532525
- DOI: 10.1152/ajplung.00192.2012
Antioxidant mimetics modulate oxidative stress and cellular signaling in airway epithelial cells infected with respiratory syncytial virus
Abstract
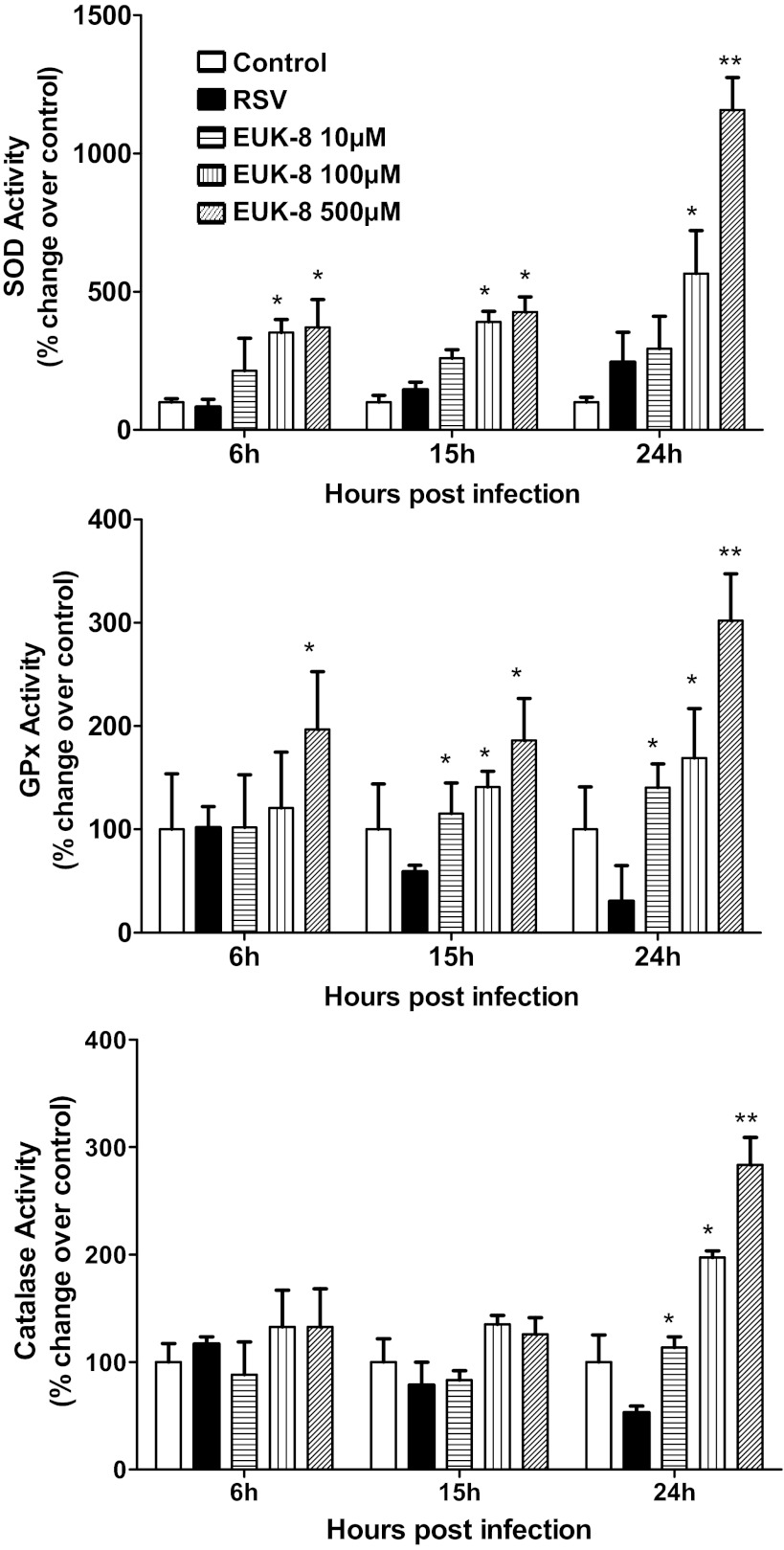
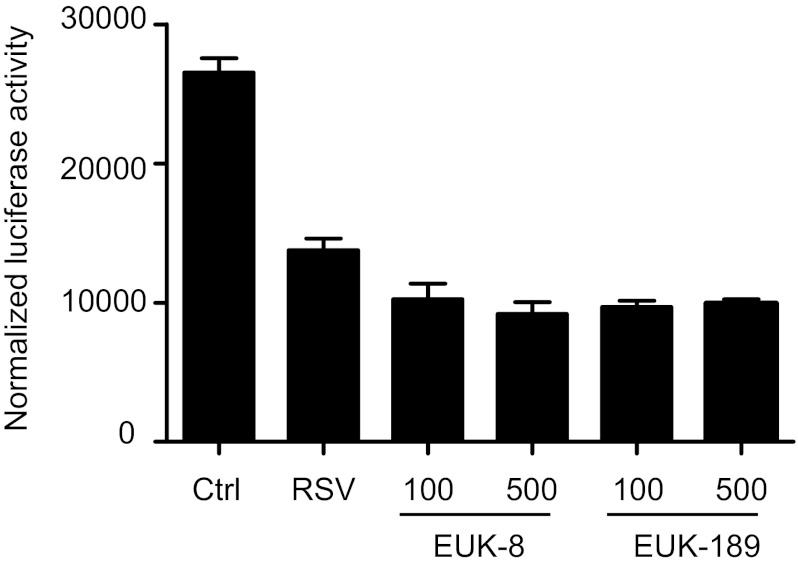
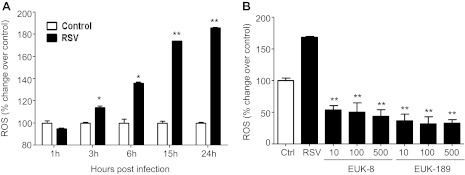
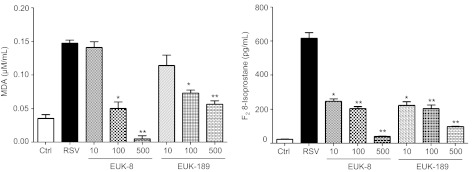
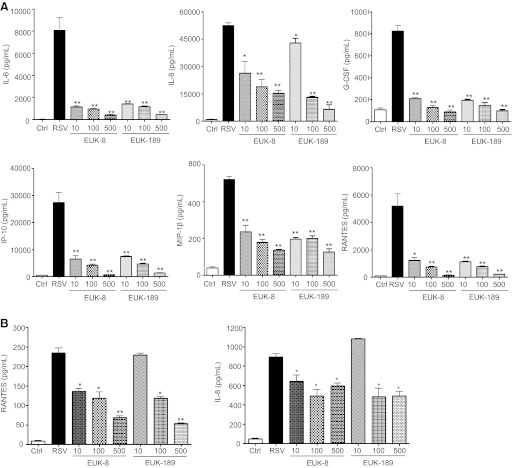
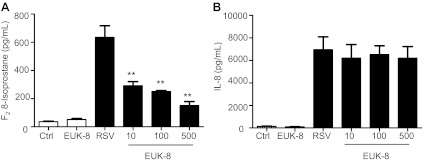
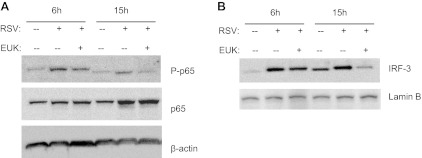
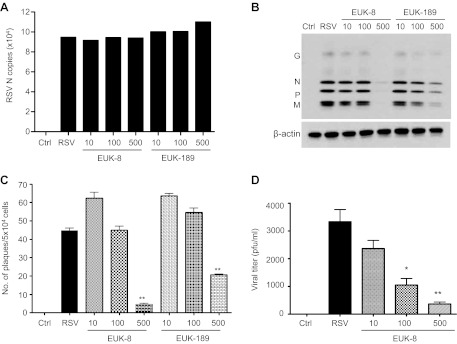
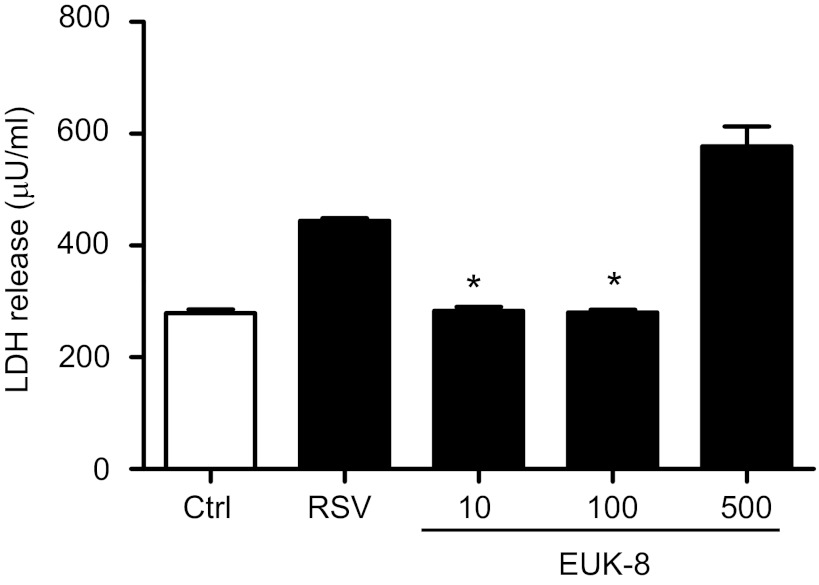
Respiratory syncytial virus (RSV) is one of the most common causes of bronchiolitis and pneumonia among infants and young children worldwide. In previous investigations, we have shown that RSV infection induces rapid generation of reactive oxygen species (ROS), which modulate viral-induced cellular signaling, and downregulation of antioxidant enzyme (AOE) expression, resulting in oxidative stress in vitro and in vivo, which plays a pathogenetic role in RSV-induced lung disease. In this study, we determined whether pharmacological intervention with synthetic catalytic scavengers could reduce RSV-induced proinflammatory gene expression and oxidative cell damage in an in vitro model of infection. Treatment of airway epithelial cells (AECs) with the salen-manganese complexes EUK-8 or EUK-189, which possess superoxide dismutase, catalase, and glutathione peroxidase activity, strongly reduced RSV-induced ROS formation by increasing cellular AOE enzymatic activity and levels of the lipid peroxidation products F(2)-8-isoprostane and malondialdehyde, which are markers of oxidative stress. Treatment of AECs with AOE mimetics also significantly inhibited RSV-induced cytokine and chemokine secretion and activation of the transcription factors nuclear factor-κB and interferon regulatory factor-3, which orchestrate proinflammatory gene expression. Both EUKs were able to reduce viral replication, when used at high doses. These results suggest that increasing antioxidant cellular capacities can significantly impact RSV-associated oxidative cell damage and cellular signaling and could represent a novel therapeutic approach in modulating virus-induced lung disease.
Figures
References
-
- Allen RG, Tresini M. Oxidative stress and gene regulation. Free Rad Biol Med 28: 463–499, 2000 - PubMed
-
- Bitko V, Velazquez A, Yank L, Yang YC, Barik S. Transcriptional induction of multiple cytokines by human respiratory syncytial virus requires activation of NF-kB and is inhibited by sodium salicylate and aspirin. Virology 232: 369–378, 1997 - PubMed
-
- Brasier AR, Spratt H, Wu Z, Boldogh I, Zhang Y, Garofal RP, Casola A, Pashmi J, Haag A, Luxon B, Kurosky A. Nuclear heat shock response and novel nuclear domain 10 reogranization in respiratory syncytial virus-infected A549 cells identified by high resolution 2D gel electrophoresis. J Virol 78: 11461–11476, 2004 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous