

AtPRK2 promotes ROP1 activation via RopGEFs in the control of polarized pollen tube growth
- PMID: 23024212
- PMCID: PMC3888354
- DOI: 10.1093/mp/sss103
AtPRK2 promotes ROP1 activation via RopGEFs in the control of polarized pollen tube growth
Abstract
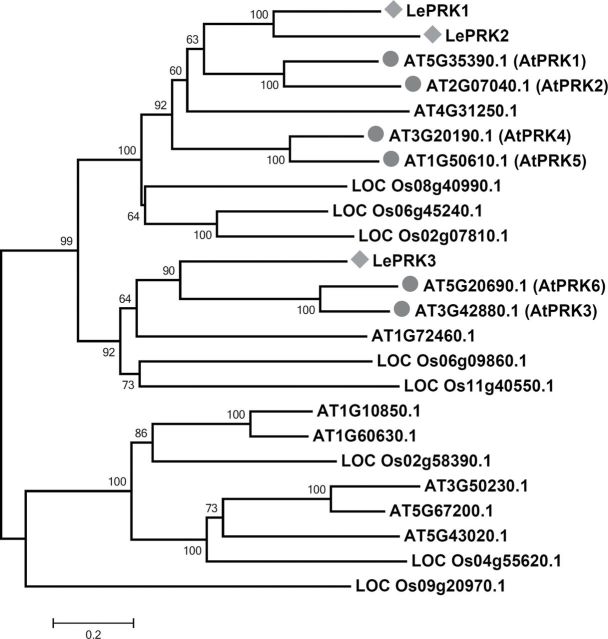
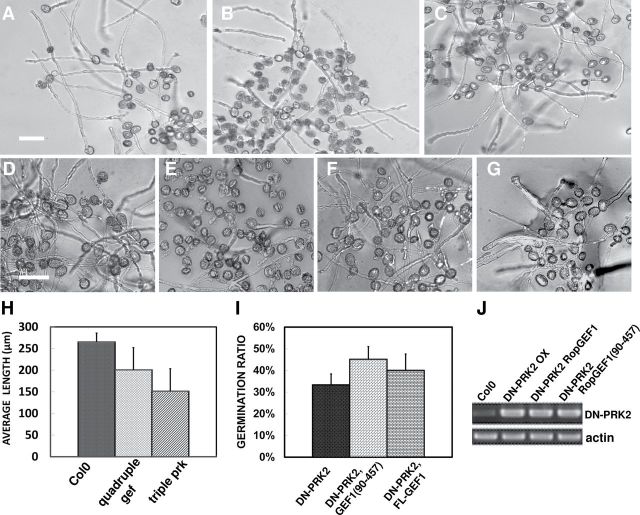
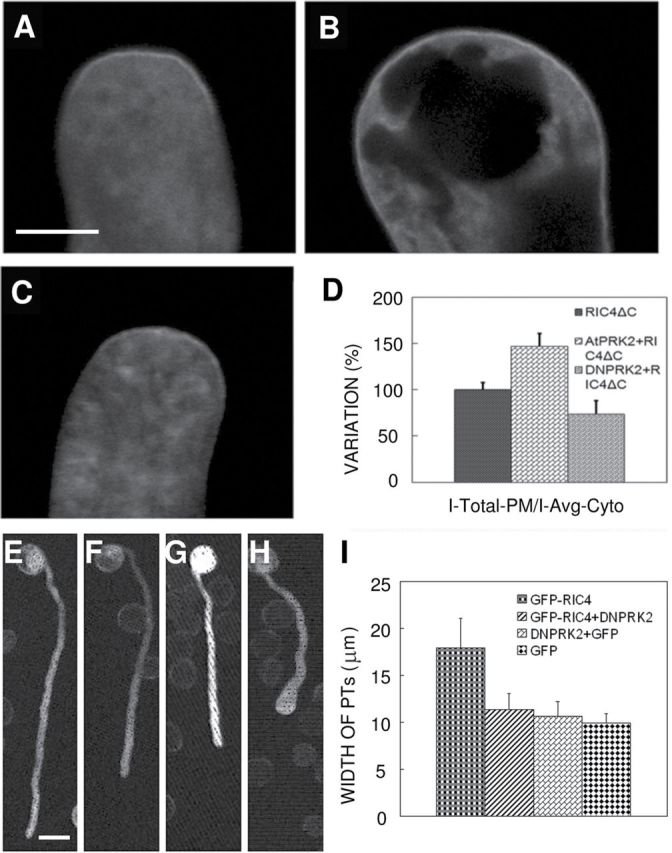
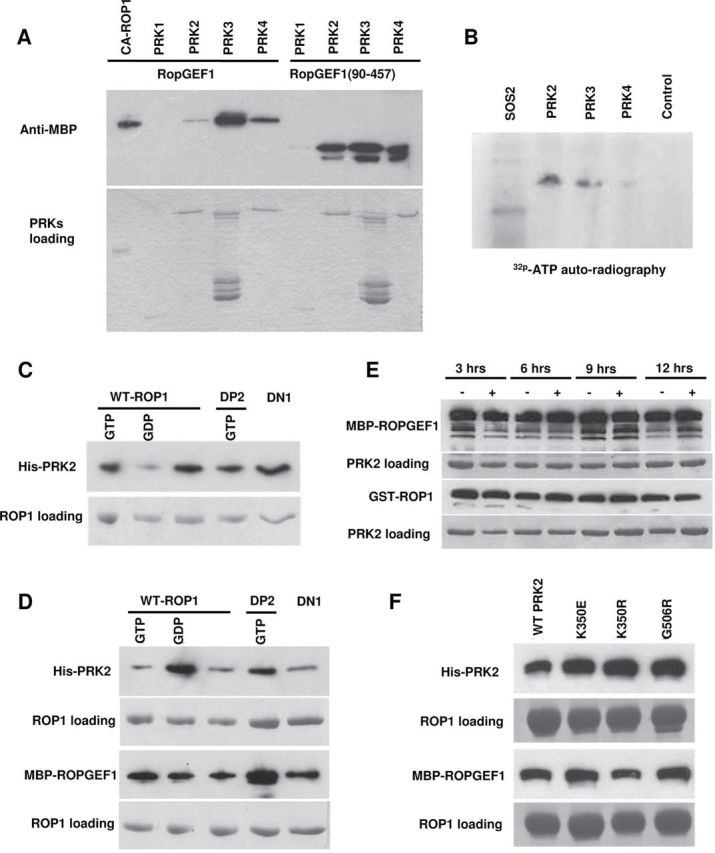
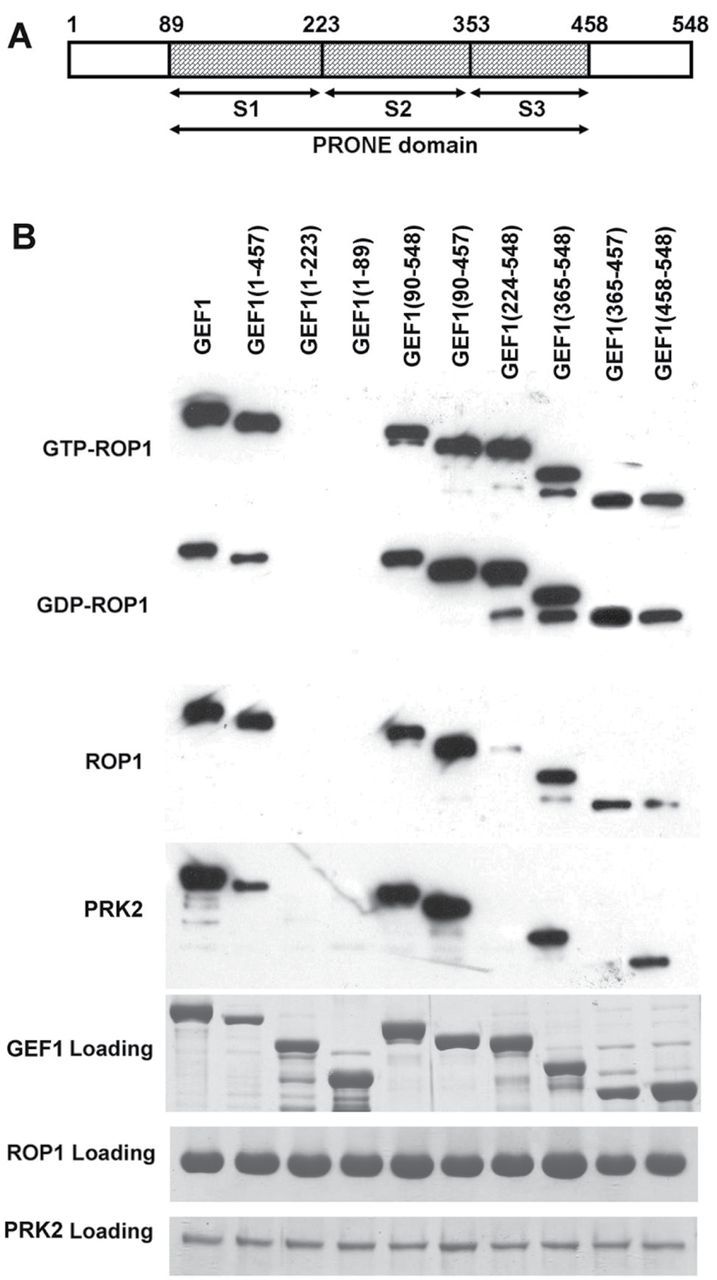
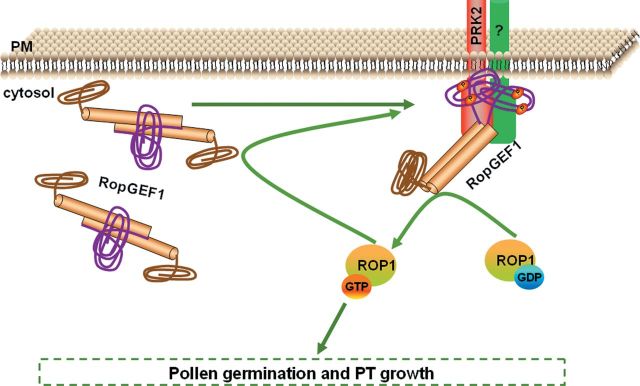
The ROP1 GTPase-based signaling network controls tip growth in Arabidopsis pollen tubes. Our previous studies imply that ROP1 might be directly activated by RopGEF1, which belongs to a plant-specific family of Rho guanine nucleotide exchange factors (RopGEFs) and in turn may be activated by an unknown factor through releasing RopGEF1's auto-inhibition. In this study, we found that RopGEF1 forms a complex with ROP1 and AtPRK2, a receptor-like protein kinase previously shown to interact with RopGEFs. AtPRK2 phosphorylated RopGEF1 in vitro and the atprk1,2,5 triple mutant showed defective pollen tube growth, similar to the phenotype of the ropgef1,9,12,14 quadruple mutant. Overexpression of a dominant negative form of AtPRK2 (DN-PRK2) inhibited pollen germination in Arabidopsis and reduced pollen elongation in tobacco. The DN-PRK2-induced pollen germination defect was rescued by overexpressing a constitutively active form of RopGEF1, RopGEF1(90-457), implying that RopGEF1 acts downstream of AtPRK2. Moreover, AtPRK2 increased ROP1 activity at the apical plasma membrane whereas DN-PRK2 reduced ROP1 activity. Finally, two mutations at the C-terminal putative phosphorylation sites of RopGEF1 (RopGEF1S460A and RopGEF1S480A) eliminated the function of RopGEF1 in vivo. Taken together, our results support the hypothesis that AtPRK2 acts as a positive regulator of the ROP1 signaling pathway most likely by activating RopGEF1 through phosphorylation.
Keywords: AtPRK2; ROP GTPase; RopGEF1; auto-inhibition; polarity growth..
Figures
References
-
- Berken A., Thomas C., Wittinghofer A. 2005. A new family of RhoGEFs activates the Rop molecular switch in plants. Nature. 436, 1176–1180 - PubMed
-
- Chang F., Yan A., Zhao L., Wu W., Yang Z. 2007. A putative calcium-permeable Cyclic Nucleotide-Gated Channel, CNGC18, regulates polarized pollen tube growth. J. Int. Plant Biol. 49, 1261–1270
-
- Cheung A.Y., Wu H.M. 2008. Structural and signaling networks for the polar cell growth machinery in pollen tubes. Annu. Rev. Plant Biol. 59, 547–572 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
