

Gene cluster encoding cholate catabolism in Rhodococcus spp
- PMID: 23024343
- PMCID: PMC3510600
- DOI: 10.1128/JB.01169-12
Gene cluster encoding cholate catabolism in Rhodococcus spp
Abstract
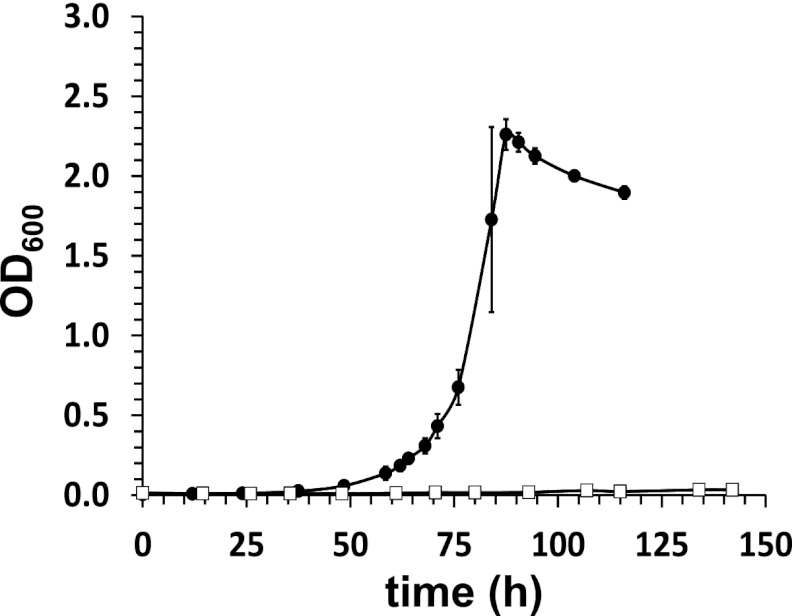
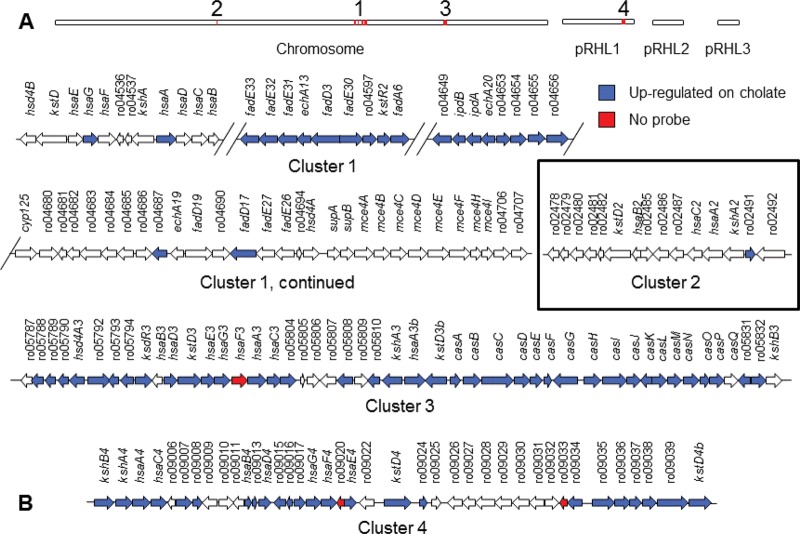
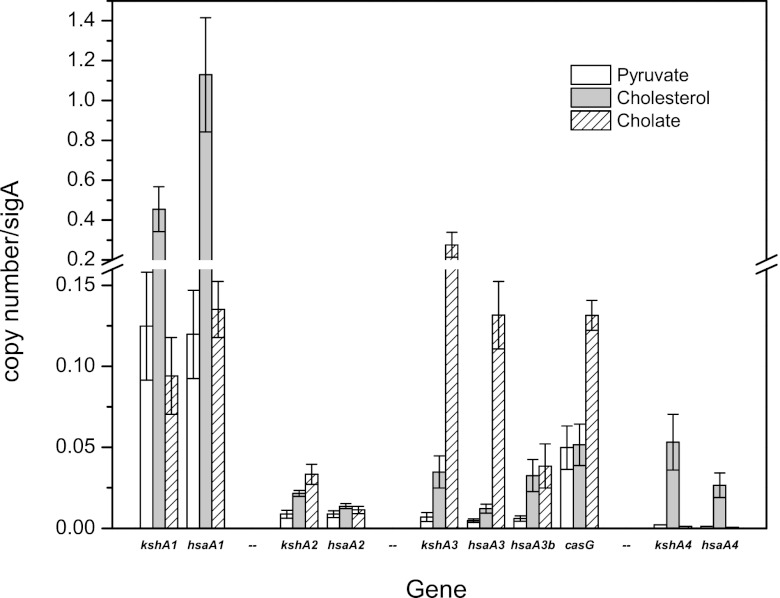
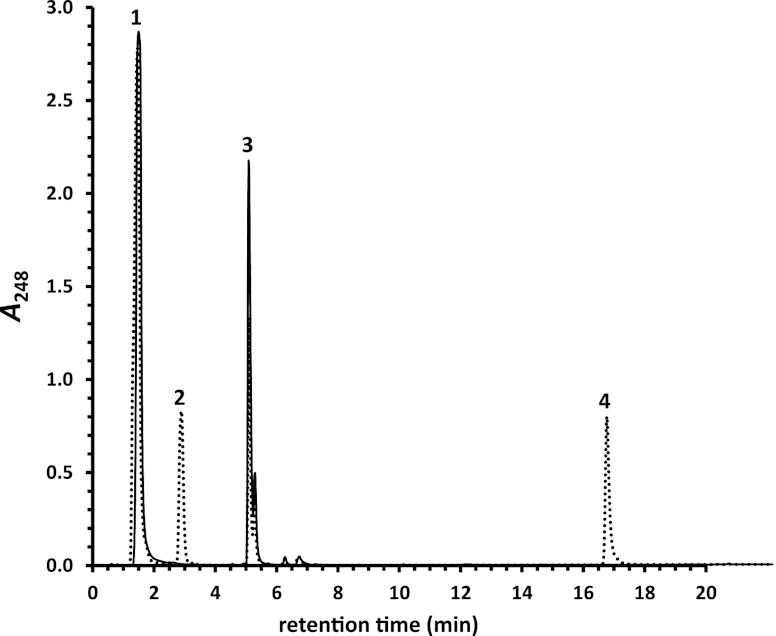
Bile acids are highly abundant steroids with important functions in vertebrate digestion. Their catabolism by bacteria is an important component of the carbon cycle, contributes to gut ecology, and has potential commercial applications. We found that Rhodococcus jostii RHA1 grows well on cholate, as well as on its conjugates, taurocholate and glycocholate. The transcriptome of RHA1 growing on cholate revealed 39 genes upregulated on cholate, occurring in a single gene cluster. Reverse transcriptase quantitative PCR confirmed that selected genes in the cluster were upregulated 10-fold on cholate versus on cholesterol. One of these genes, kshA3, encoding a putative 3-ketosteroid-9α-hydroxylase, was deleted and found essential for growth on cholate. Two coenzyme A (CoA) synthetases encoded in the cluster, CasG and CasI, were heterologously expressed. CasG was shown to transform cholate to cholyl-CoA, thus initiating side chain degradation. CasI was shown to form CoA derivatives of steroids with isopropanoyl side chains, likely occurring as degradation intermediates. Orthologous gene clusters were identified in all available Rhodococcus genomes, as well as that of Thermomonospora curvata. Moreover, Rhodococcus equi 103S, Rhodococcus ruber Chol-4 and Rhodococcus erythropolis SQ1 each grew on cholate. In contrast, several mycolic acid bacteria lacking the gene cluster were unable to grow on cholate. Our results demonstrate that the above-mentioned gene cluster encodes cholate catabolism and is distinct from a more widely occurring gene cluster encoding cholesterol catabolism.
Figures
References
-
- Achterholt S, Priefert H, Steinbuchel A. 2000. Identification of Amycolatopsis sp. strain HR167 genes, involved in the bioconversion of ferulic acid to vanillin. Appl. Microbiol. Biotechnol. 54:799–807 - PubMed
-
- Birkenmaier A, Möller HM, Philipp B. 2011. Identification of a thiolase gene essential for β-oxidation of the acyl side chain of the steroid compound cholate in Pseudomonas sp. strain Chol1. FEMS Microbiol. Lett. 318:123–130 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
