

A critical reevaluation of the stationary axonal cytoskeleton hypothesis
- PMID: 23027591
- PMCID: PMC3725768
- DOI: 10.1002/cm.21083
A critical reevaluation of the stationary axonal cytoskeleton hypothesis
Abstract
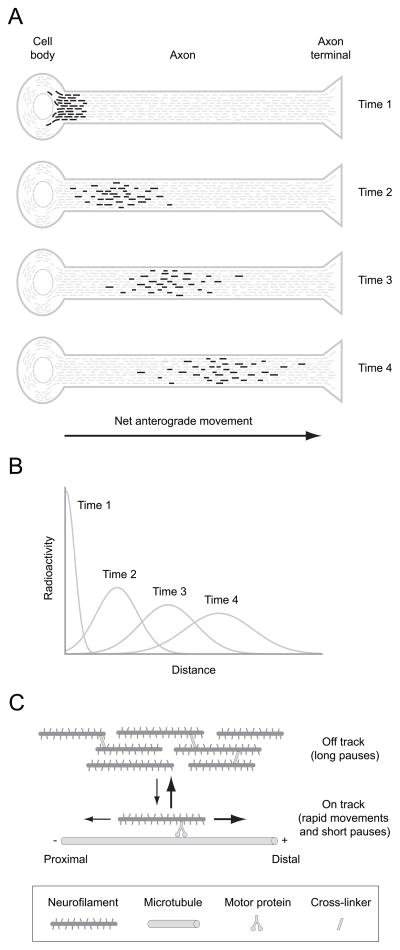
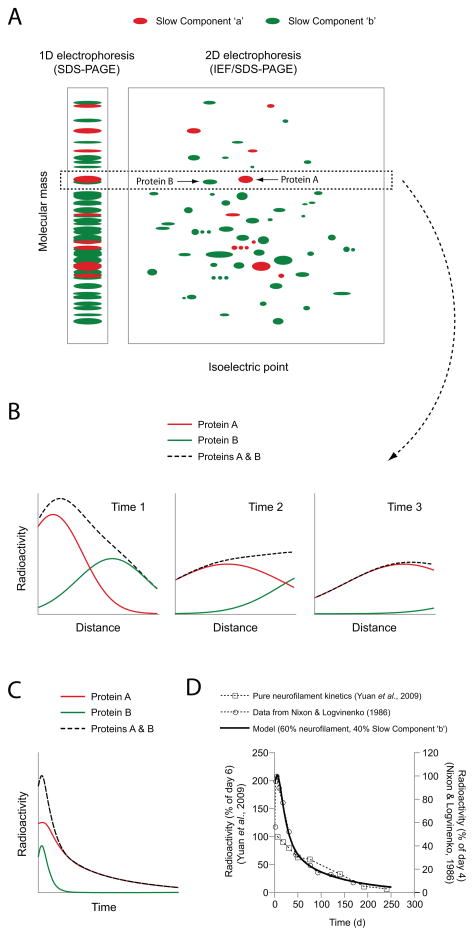
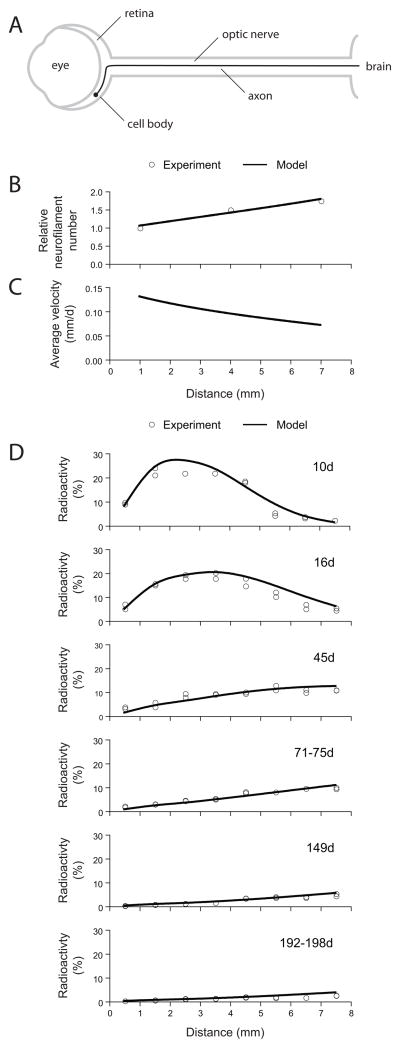
Neurofilaments are transported along axons in a rapid intermittent and bidirectional manner but there is a long-standing controversy about whether this applies to all axonal neurofilaments. Some have proposed that only a small proportion of axonal neurofilaments are mobile and that most are deposited into a persistently stationary and extensively cross-linked cytoskeleton that remains fixed in place for many months without movement, turning over very slowly. In contrast, others have proposed that this hypothesis is based on a misinterpretation of the experimental data and that, in fact, all axonal neurofilaments move. These contrary perspectives have distinct implications for our understanding of how neurofilaments are organized and reorganized in axons both in health and disease. Here, we discuss the history and substance of this controversy. We show that the published data on the kinetics and distribution of neurofilaments along axons favor a simple "stop and go" transport model in which axons contain a single population of neurofilaments that all move in a stochastic, bidirectional and intermittent manner. Based on these considerations, we propose a dynamic view of the neuronal cytoskeleton in which all neurofilaments cycle repeatedly between moving and pausing states throughout their journey along the axon. The filaments move infrequently, but the average pause duration is on the order of hours rather than weeks or months. Against this fluid backdrop, the action of molecular motors on neurofilaments can have dramatic effects on neurofilament organization that would not be possible if the neurofilaments were extensively cross-linked into a truly stationary network.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
References
-
- Baas PW, Brown A. Slow axonal transport: the polymer transport model. Trends in Cell Biology. 1997;7(10):380–384. - PubMed
-
- Barry DM, Millecamps S, Julien JP, Garcia ML. New movements in neurofilament transport, turnover and disease. Experimental Cell Research. 2007;313(10):2110–20. - PubMed
-
- Black MM, Lasek RJ. Axonal transport of actin: slow component b is the principal source of actin for the axon. Brain Research. 1979;171(3):401–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
