

Chlamydia trachomatis protein GrgA activates transcription by contacting the nonconserved region of σ66
- PMID: 23027952
- PMCID: PMC3479454
- DOI: 10.1073/pnas.1207300109
Chlamydia trachomatis protein GrgA activates transcription by contacting the nonconserved region of σ66
Abstract
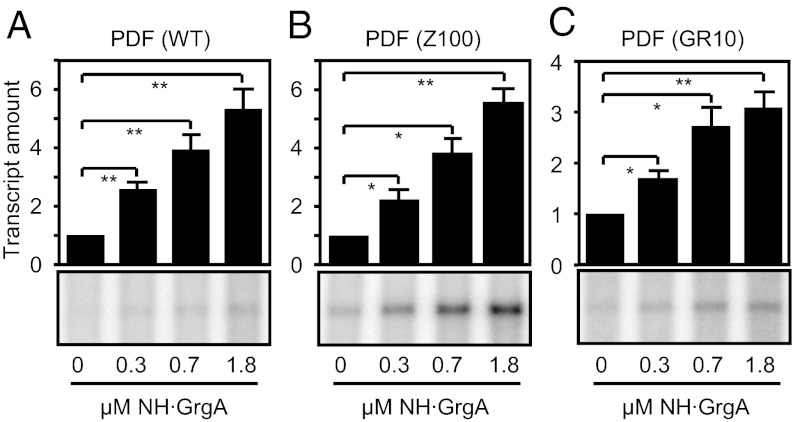
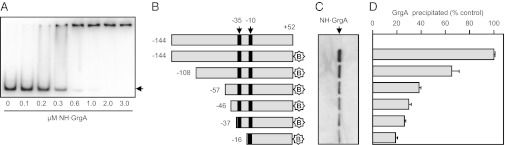
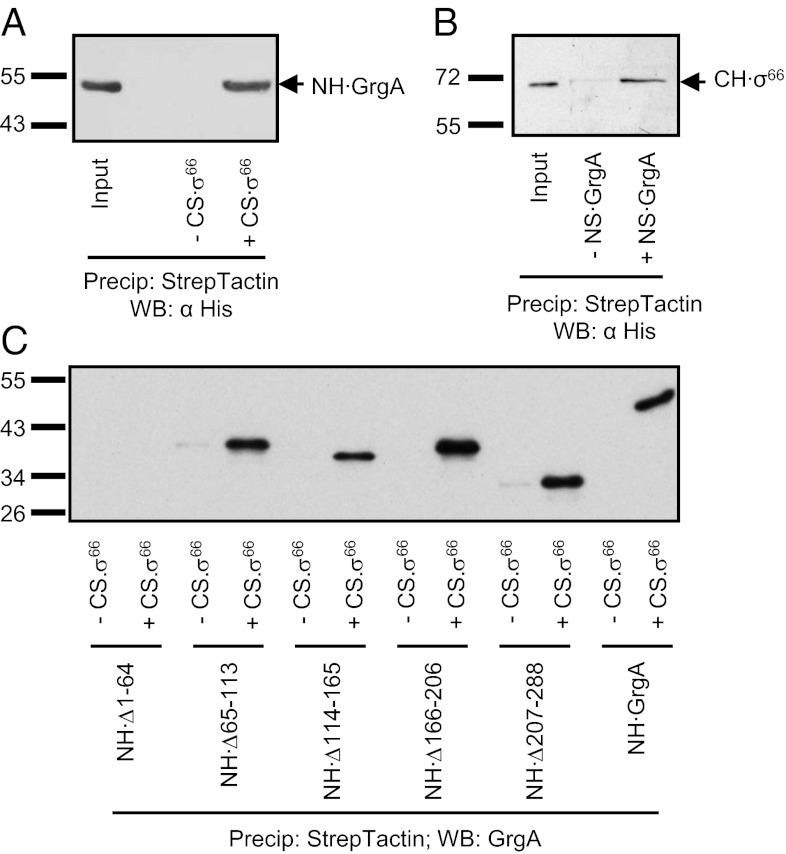
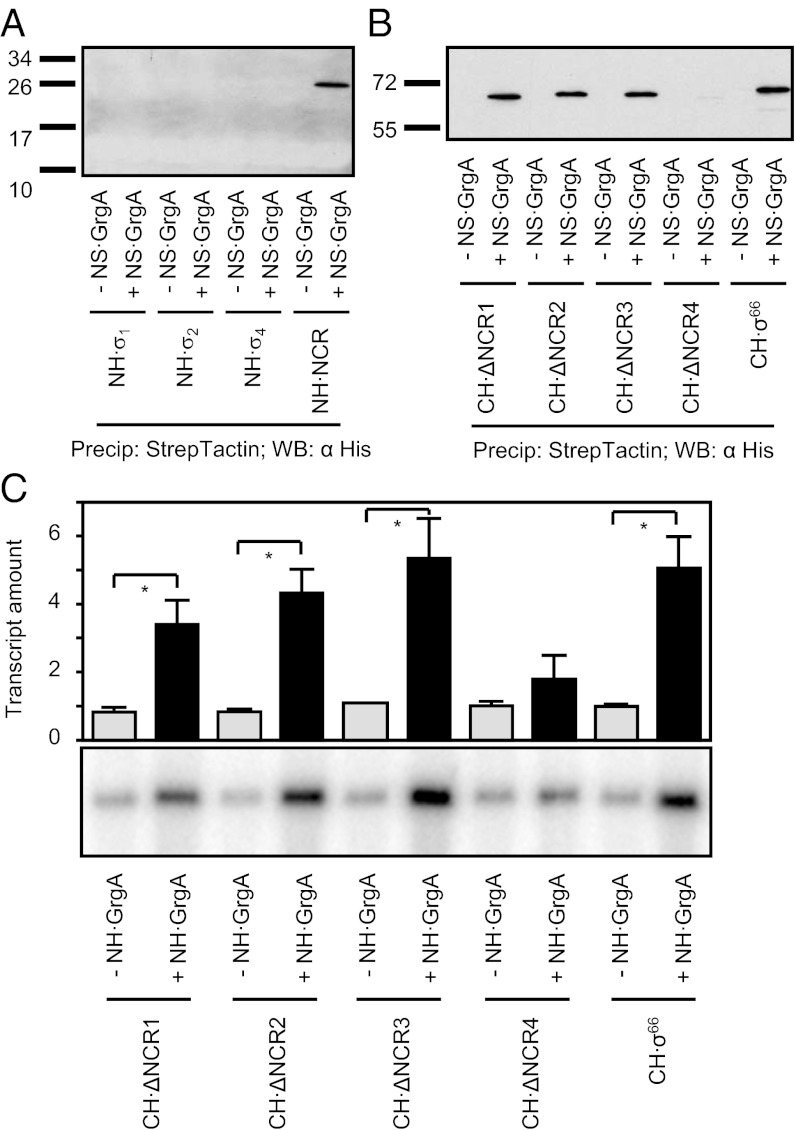
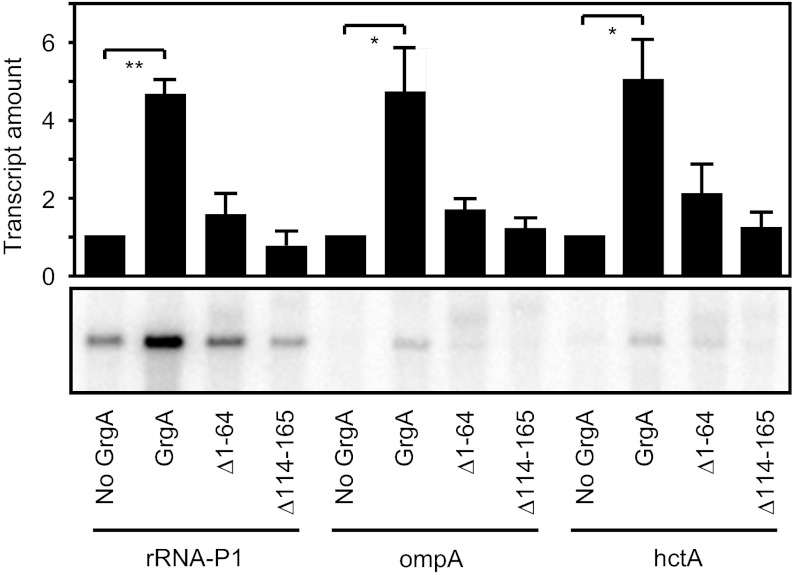
The bacterial RNA polymerase holoenzyme consists of a catalytic core enzyme in complex with a σ factor that is required for promoter-specific transcription initiation. Primary, or housekeeping, σ factors are responsible for most of the gene expression that occurs during the exponential phase of growth. Primary σ factors share four regions of conserved sequence, regions 1-4, which have been further subdivided. Many primary σ factors also contain a nonconserved region (NCR) located between subregions 1.2 and 2.1, which can vary widely in length. Interactions between the NCR of the primary σ factor of Escherichia coli, σ(70), and the β' subunit of the E. coli core enzyme have been shown to influence gene expression, suggesting that the NCR of primary σ factors represents a potential target for transcription regulation. Here, we report the identification and characterization of a previously undocumented Chlamydia trachomatis transcription factor, designated GrgA (general regulator of genes A). We demonstrate in vitro that GrgA is a DNA-binding protein that can stimulate transcription from a range of σ(66)-dependent promoters. We further show that GrgA activates transcription by contacting the NCR of the primary σ factor of C. trachomatis, σ(66). Our findings suggest GrgA serves as an important regulator of σ(66)-dependent transcription in C. trachomatis. Furthermore, because GrgA is present only in chlamydiae, our findings highlight how nonconserved regions of the bacterial RNA polymerase can be targets of regulatory factors that are unique to particular organisms.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Stephens RS, Myers G, Eppinger M, Bavoil PM. Divergence without difference: Phylogenetics and taxonomy of Chlamydia resolved. FEMS Immunol Med Microbiol. 2009;55:115–119. - PubMed
-
- Schachter J. Infection and disease epidemiology. In: Stephens RS, editor. Chlamydia Intracellular Biology, Pathogenesis. Washington, DC: ASM Press; 1999. pp. 139–169.
-
- Todd WJ, Caldwell HD. The interaction of Chlamydia trachomatis with host cells: Ultrastructural studies of the mechanism of release of a biovar II strain from HeLa 229 cells. J Infect Dis. 1985;151:1037–1044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
