

Synaptic vesicle clustering requires a distinct MIG-10/Lamellipodin isoform and ABI-1 downstream from Netrin
- PMID: 23028145
- PMCID: PMC3465741
- DOI: 10.1101/gad.193409.112
Synaptic vesicle clustering requires a distinct MIG-10/Lamellipodin isoform and ABI-1 downstream from Netrin
Abstract
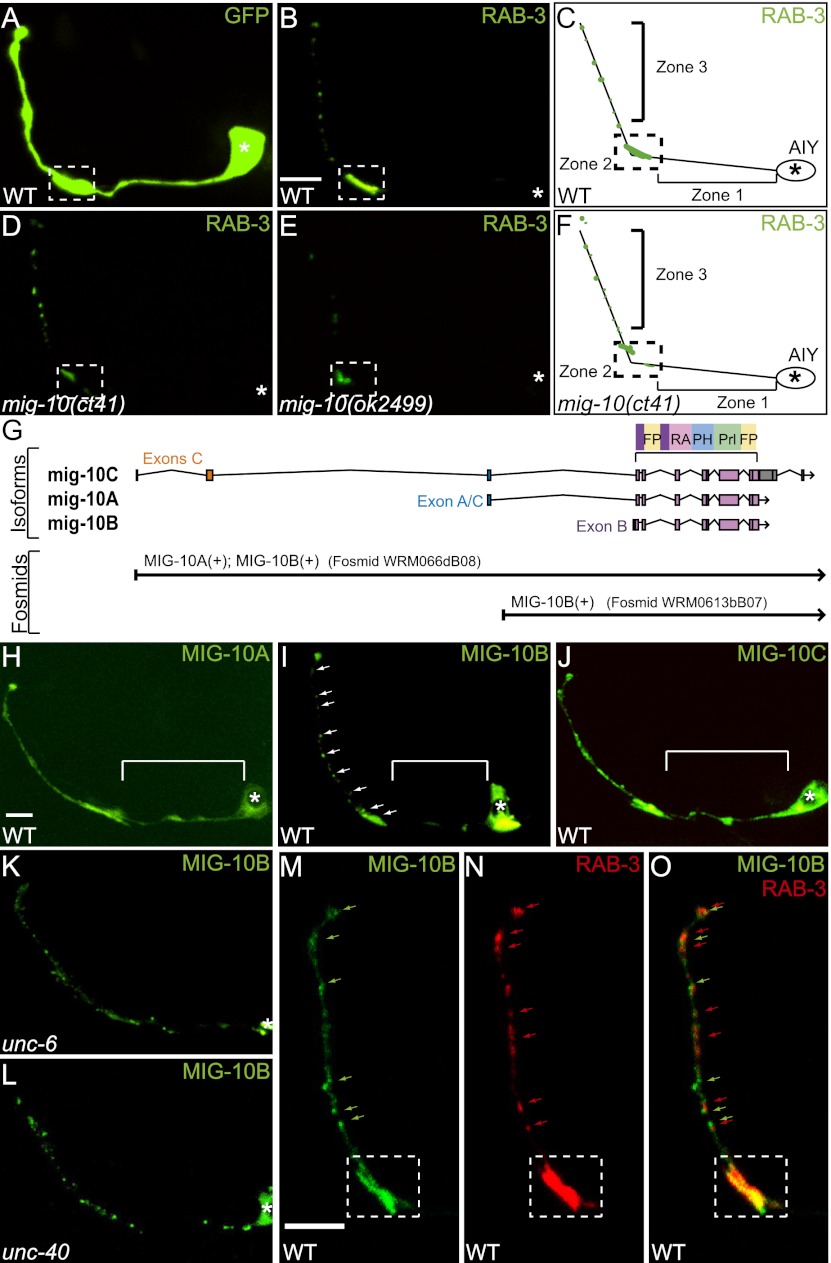
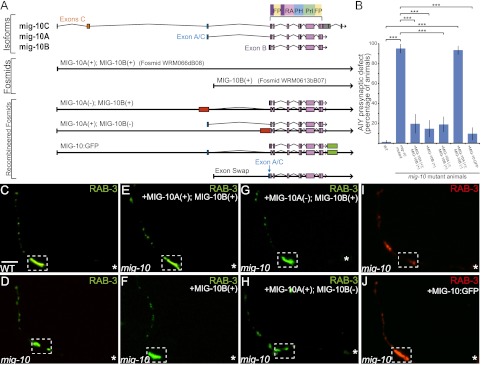
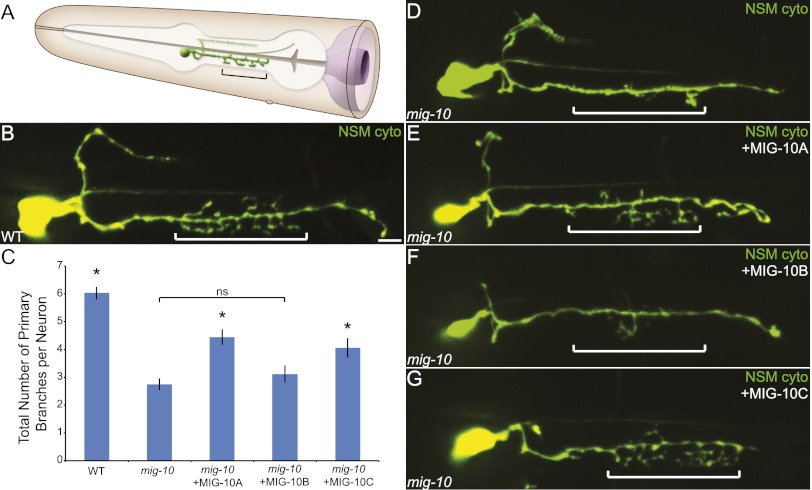
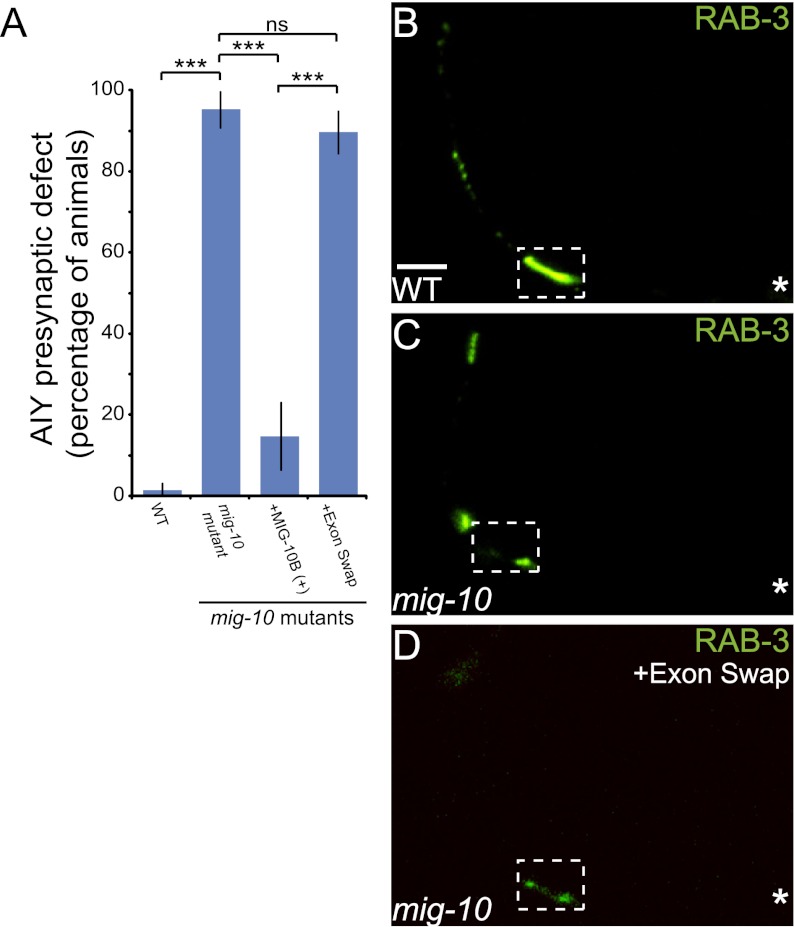
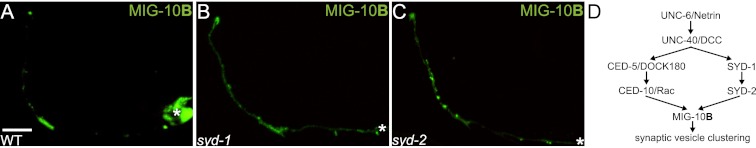
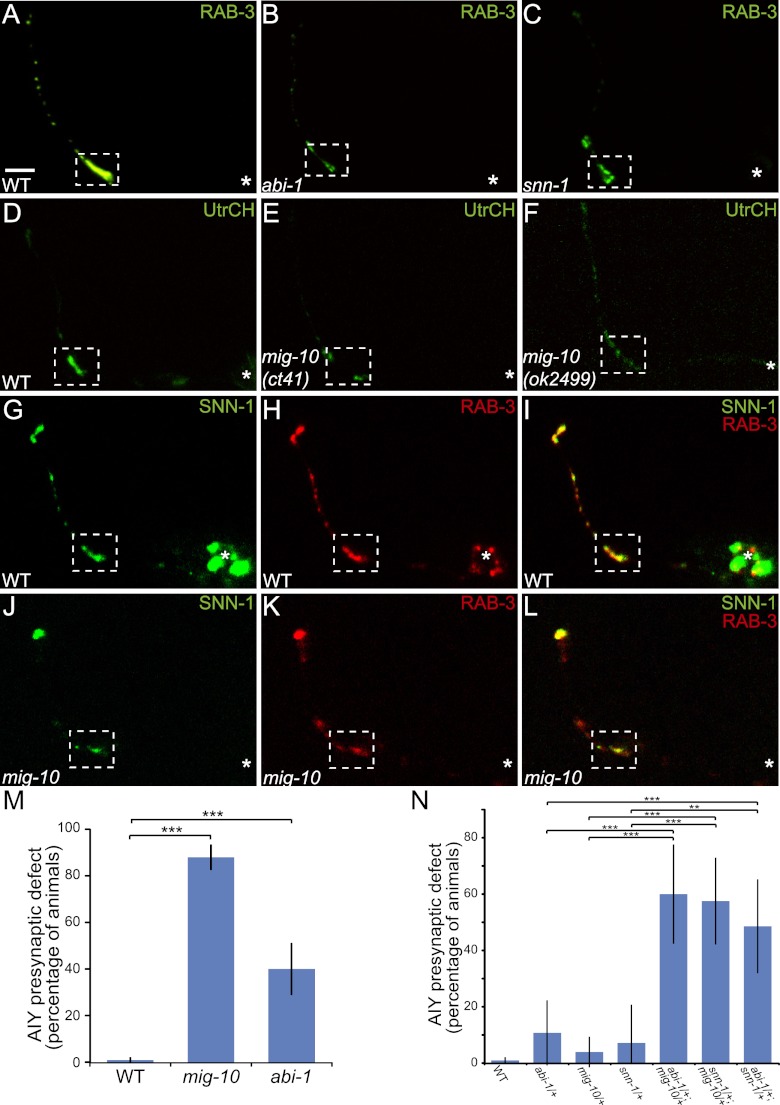
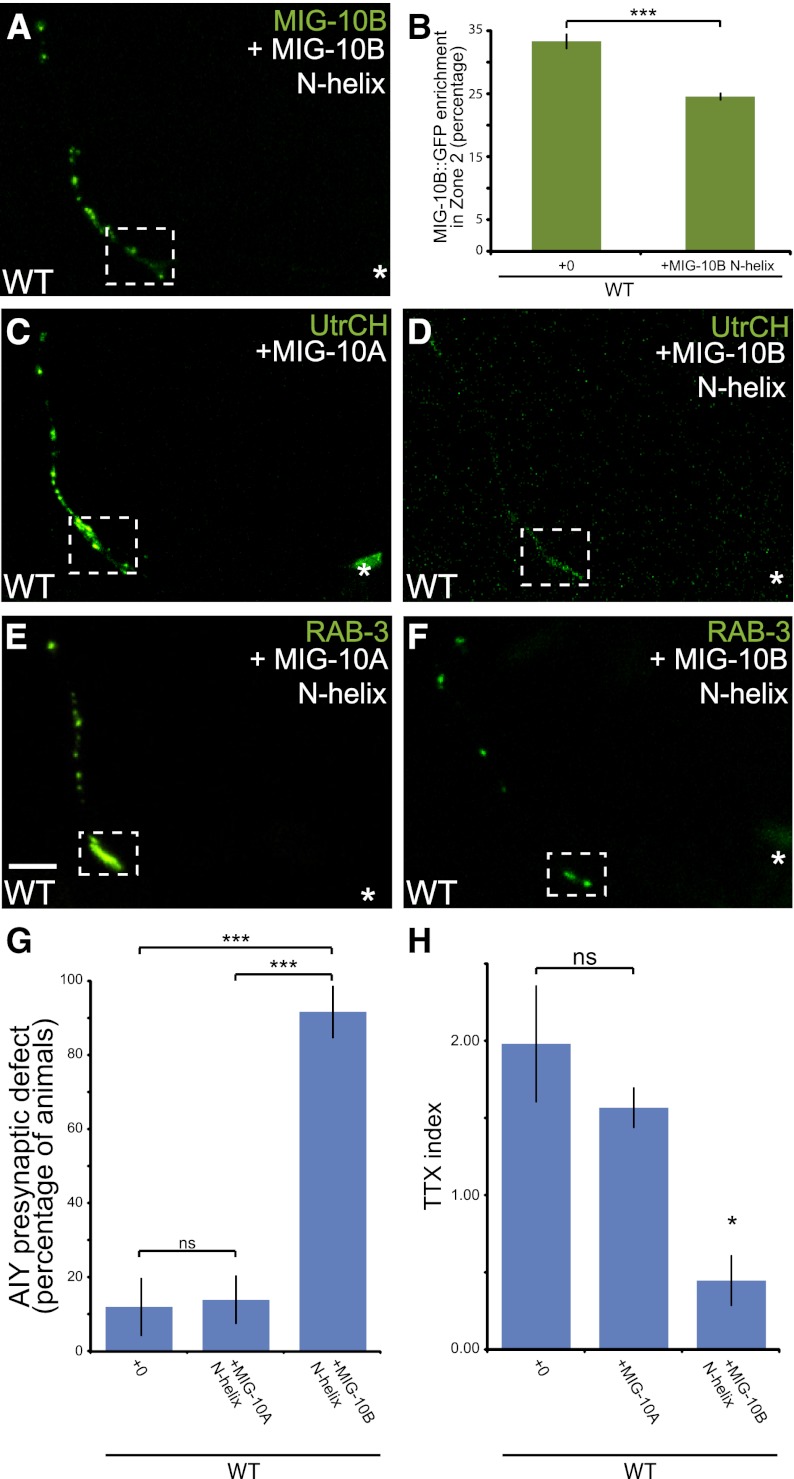
The chemotrophic factor Netrin can simultaneously instruct different neurodevelopmental programs in individual neurons in vivo. How neurons correctly interpret the Netrin signal and undergo the appropriate neurodevelopmental response is not understood. Here we identify MIG-10 isoforms as critical determinants of individual cellular responses to Netrin. We determined that distinct MIG-10 isoforms, varying only in their N-terminal motifs, can localize to specific subcellular domains and are differentially required for discrete neurodevelopmental processes in vivo. We identified MIG-10B as an isoform uniquely capable of localizing to presynaptic regions and instructing synaptic vesicle clustering in response to Netrin. MIG-10B interacts with Abl-interacting protein-1 (ABI-1)/Abi1, a component of the WAVE complex, to organize the actin cytoskeleton at presynaptic sites and instruct vesicle clustering through SNN-1/Synapsin. We identified a motif in the MIG-10B N-terminal domain that is required for its function and localization to presynaptic sites. With this motif, we engineered a dominant-negative MIG-10B construct that disrupts vesicle clustering and animal thermotaxis behavior when expressed in a single neuron in vivo. Our findings indicate that the unique N-terminal domains confer distinct MIG-10 isoforms with unique capabilities to localize to distinct subcellular compartments, organize the actin cytoskeleton at these sites, and instruct distinct Netrin-dependent neurodevelopmental programs.
Figures
Similar articles
-
MIG-10 (lamellipodin) has netrin-independent functions and is a FOS-1A transcriptional target during anchor cell invasion in C. elegans.Development. 2014 Mar;141(6):1342-53. doi: 10.1242/dev.102434. Epub 2014 Feb 19. Development. 2014. PMID: 24553288 Free PMC article.
-
Netrin instructs synaptic vesicle clustering through Rac GTPase, MIG-10, and the actin cytoskeleton.J Cell Biol. 2012 Apr 2;197(1):75-88. doi: 10.1083/jcb.201110127. Epub 2012 Mar 26. J Cell Biol. 2012. PMID: 22451697 Free PMC article.
-
Abelson interactor-1 (ABI-1) interacts with MRL adaptor protein MIG-10 and is required in guided cell migrations and process outgrowth in C. elegans.Dev Biol. 2013 Jan 1;373(1):1-13. doi: 10.1016/j.ydbio.2012.09.017. Epub 2012 Sep 27. Dev Biol. 2013. PMID: 23022657 Free PMC article.
-
Localization mechanisms of the axon guidance molecule UNC-6/Netrin and its receptors, UNC-5 and UNC-40, in Caenorhabditis elegans.Dev Growth Differ. 2012 Apr;54(3):390-7. doi: 10.1111/j.1440-169X.2012.01349.x. Dev Growth Differ. 2012. PMID: 22524608 Review.
-
Assembly and tissue functions of early embryonic laminins and netrins.Curr Opin Cell Biol. 2004 Oct;16(5):572-9. doi: 10.1016/j.ceb.2004.07.013. Curr Opin Cell Biol. 2004. PMID: 15363809 Review.
Cited by
-
The actin cytoskeleton in presynaptic assembly.Cell Adh Migr. 2013 Jul-Aug;7(4):379-87. doi: 10.4161/cam.24803. Epub 2013 Apr 29. Cell Adh Migr. 2013. PMID: 23628914 Free PMC article. Review.
-
MIG-10 (lamellipodin) has netrin-independent functions and is a FOS-1A transcriptional target during anchor cell invasion in C. elegans.Development. 2014 Mar;141(6):1342-53. doi: 10.1242/dev.102434. Epub 2014 Feb 19. Development. 2014. PMID: 24553288 Free PMC article.
-
The WAVE regulatory complex links diverse receptors to the actin cytoskeleton.Cell. 2014 Jan 16;156(1-2):195-207. doi: 10.1016/j.cell.2013.11.048. Cell. 2014. PMID: 24439376 Free PMC article.
-
The Role of c-Abl Tyrosine Kinase in Brain and Its Pathologies.Cells. 2023 Aug 10;12(16):2041. doi: 10.3390/cells12162041. Cells. 2023. PMID: 37626851 Free PMC article. Review.
-
SYD-1C, UNC-40 (DCC) and SAX-3 (Robo) function interdependently to promote axon guidance by regulating the MIG-2 GTPase.PLoS Genet. 2015 Apr 15;11(4):e1005185. doi: 10.1371/journal.pgen.1005185. eCollection 2015 Apr. PLoS Genet. 2015. PMID: 25876065 Free PMC article.
References
-
- Asakura T, Ogura K, Goshima Y 2007. UNC-6 expression by the vulval precursor cells of Caenorhabditis elegans is required for the complex axon guidance of the HSN neurons. Dev Biol 304: 800–810 - PubMed
-
- Bahler M, Greengard P 1987. Synapsin I bundles F-actin in a phosphorylation-dependent manner. Nature 326: 704–707 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous