

Characterization of inducible models of Tay-Sachs and related disease
- PMID: 23028353
- PMCID: PMC3447966
- DOI: 10.1371/journal.pgen.1002943
Characterization of inducible models of Tay-Sachs and related disease
Abstract
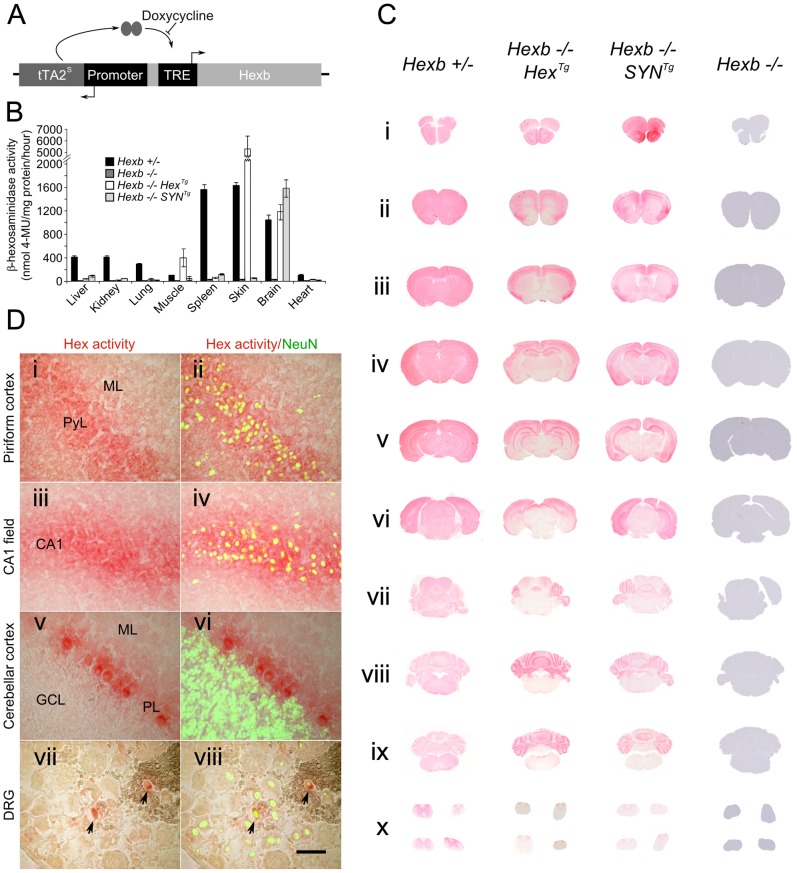
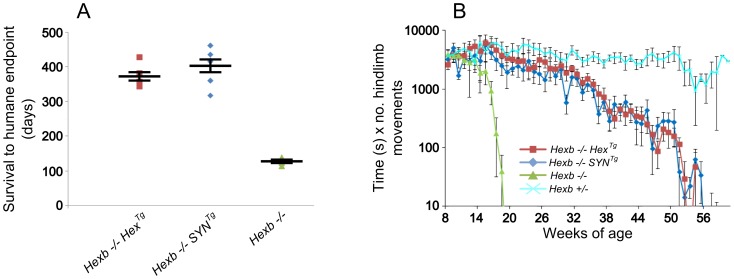
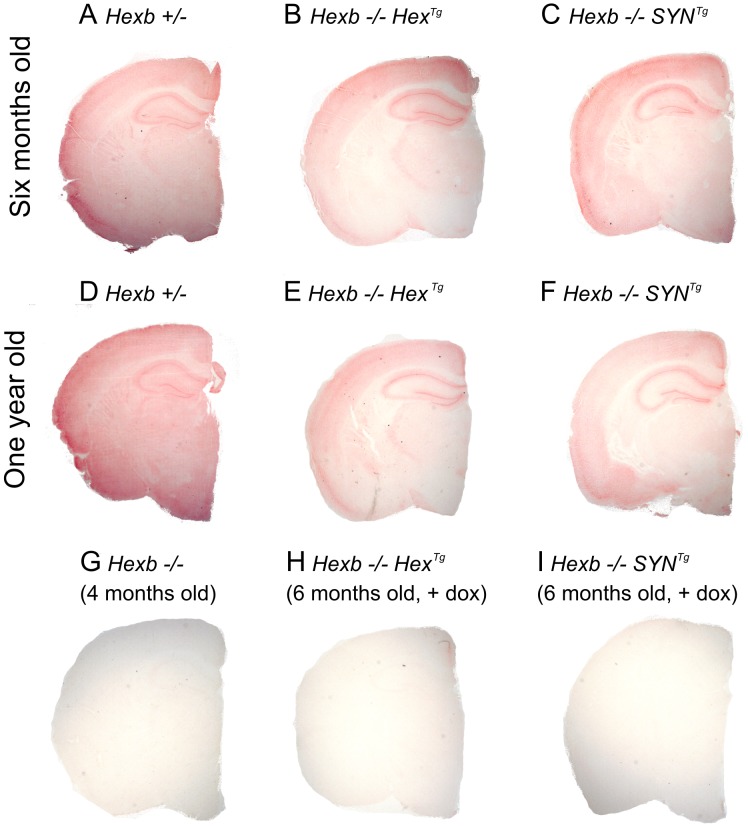
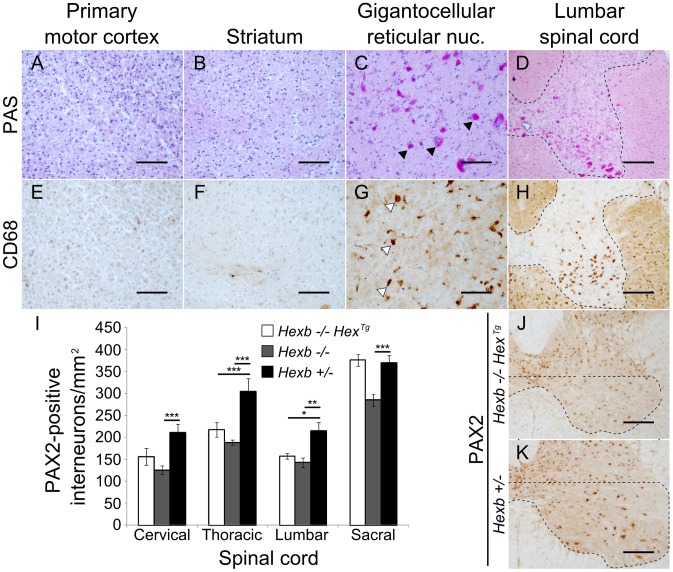
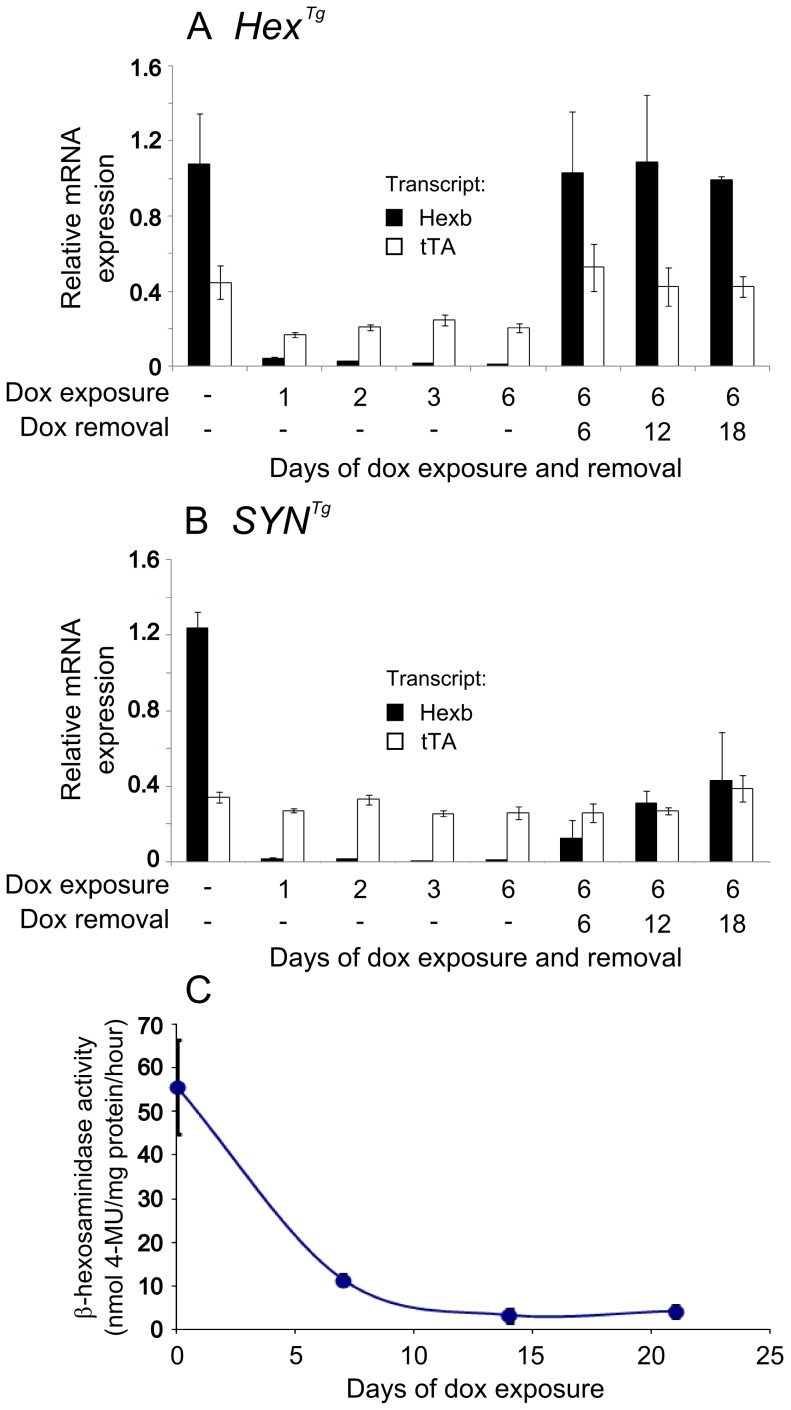
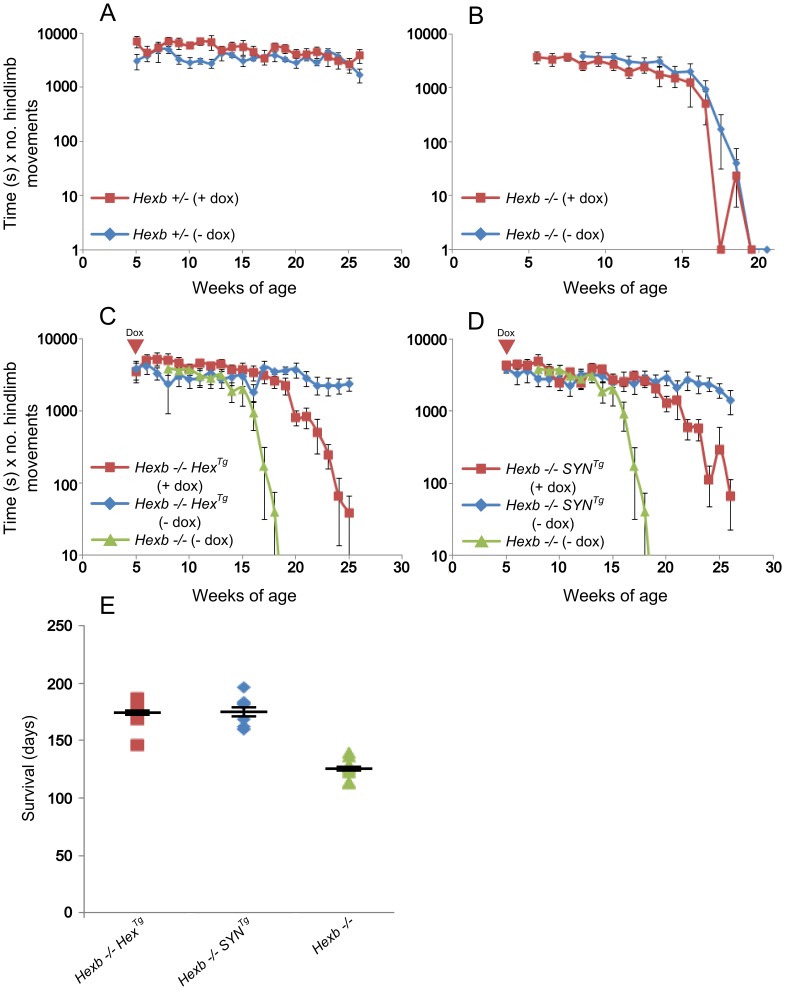
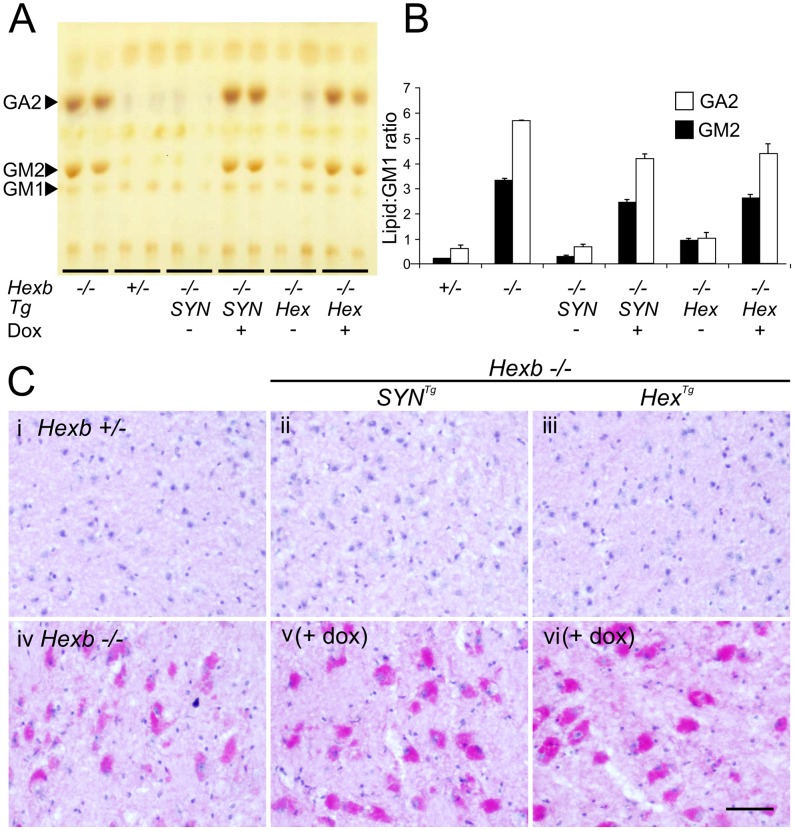
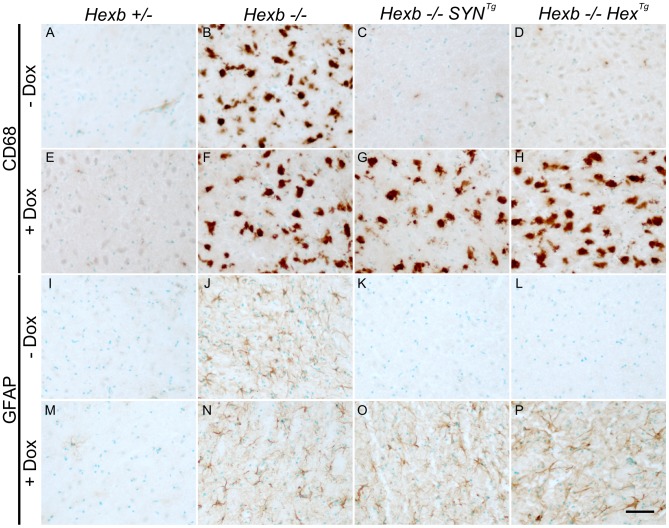
Tay-Sachs and Sandhoff diseases are lethal inborn errors of acid β-N-acetylhexosaminidase activity, characterized by lysosomal storage of GM2 ganglioside and related glycoconjugates in the nervous system. The molecular events that lead to irreversible neuronal injury accompanied by gliosis are unknown; but gene transfer, when undertaken before neurological signs are manifest, effectively rescues the acute neurodegenerative illness in Hexb-/- (Sandhoff) mice that lack β-hexosaminidases A and B. To define determinants of therapeutic efficacy and establish a dynamic experimental platform to systematically investigate cellular pathogenesis of GM2 gangliosidosis, we generated two inducible experimental models. Reversible transgenic expression of β-hexosaminidase directed by two promoters, mouse Hexb and human Synapsin 1 promoters, permitted progression of GM2 gangliosidosis in Sandhoff mice to be modified at pre-defined ages. A single auto-regulatory tetracycline-sensitive expression cassette controlled expression of transgenic Hexb in the brain of Hexb-/- mice and provided long-term rescue from the acute neuronopathic disorder, as well as the accompanying pathological storage of glycoconjugates and gliosis in most parts of the brain. Ultimately, late-onset brainstem and ventral spinal cord pathology occurred and was associated with increased tone in the limbs. Silencing transgenic Hexb expression in five-week-old mice induced stereotypic signs and progression of Sandhoff disease, including tremor, bradykinesia, and hind-limb paralysis. As in germline Hexb-/- mice, these neurodegenerative manifestations advanced rapidly, indicating that the pathogenesis and progression of GM2 gangliosidosis is not influenced by developmental events in the maturing nervous system.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Tay W (1881) Symmetrical changes in the region of the yellow spot in each eye of an infant. Trans Opthalmol Soc 1: 55–57.
-
- Sachs B (1887) On arrested cerebral development with special reference to cortical pathology. J Nerv Ment Dis 14: 541–554.
-
- Sandhoff K, Andreae U, Jatzkewitz H (1968) Deficient hexosaminidase activity in an exceptional case of Tay-Sachs disease with additional storage of kidney globoside in visceral organs. Life Sci 7: 283–288. - PubMed
-
- Svennerholm L (1962) The chemical structure of normal human brain and Tay-Sachs gangliosides. Biochem Biophys Res Commun 9: 436–441. - PubMed
-
- Makita A, Yamakawa T (1963) The glycolipids of the brain of Tay-Sachs' disease. The chemical structures of globoside and main ganglioside. Jpn J Exp Med 33: 361–368. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
