

Role of transposon-derived small RNAs in the interplay between genomes and parasitic DNA in rice
- PMID: 23028360
- PMCID: PMC3459959
- DOI: 10.1371/journal.pgen.1002953
Role of transposon-derived small RNAs in the interplay between genomes and parasitic DNA in rice
Abstract
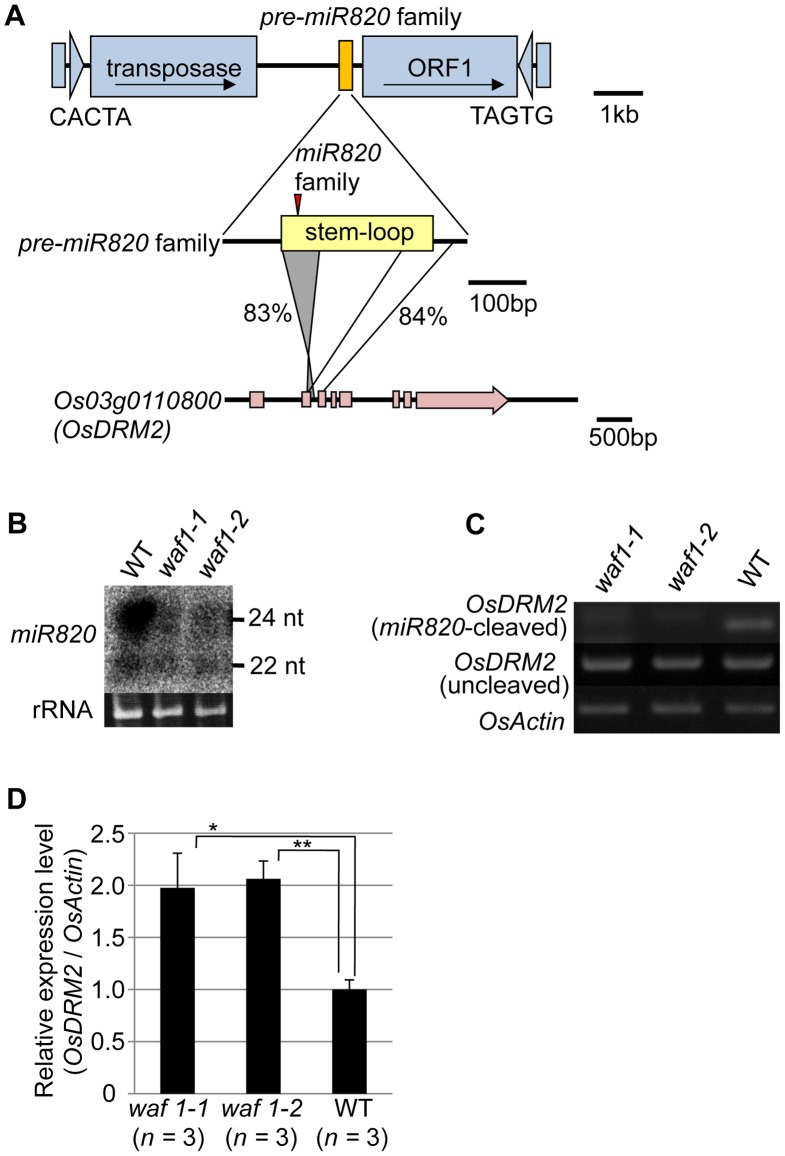
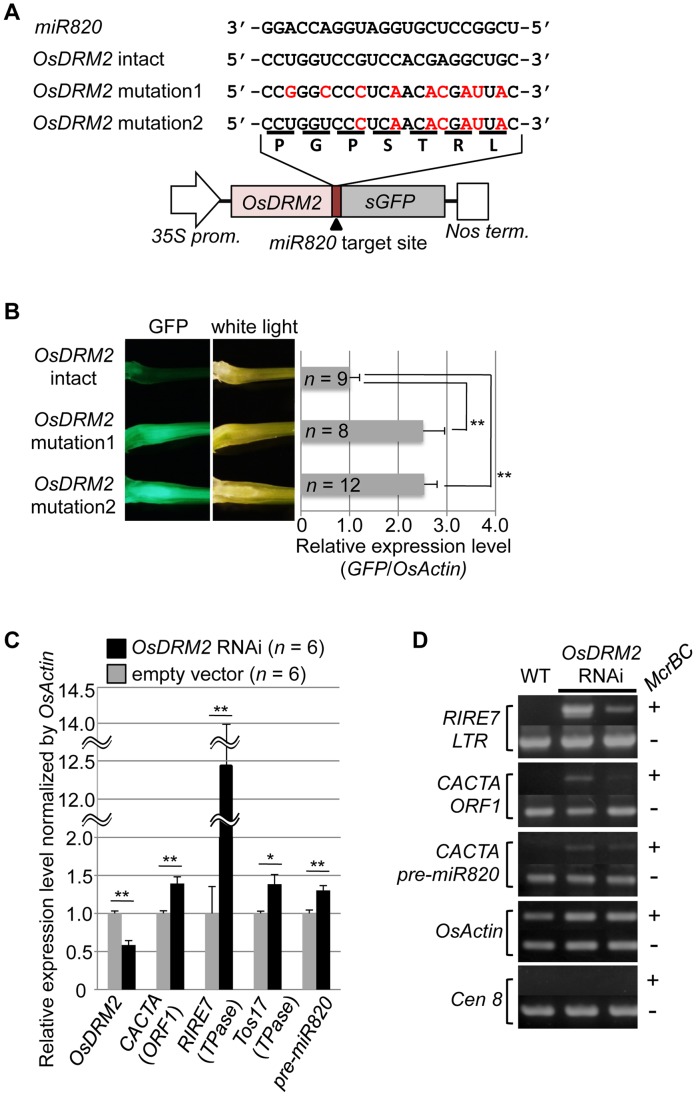
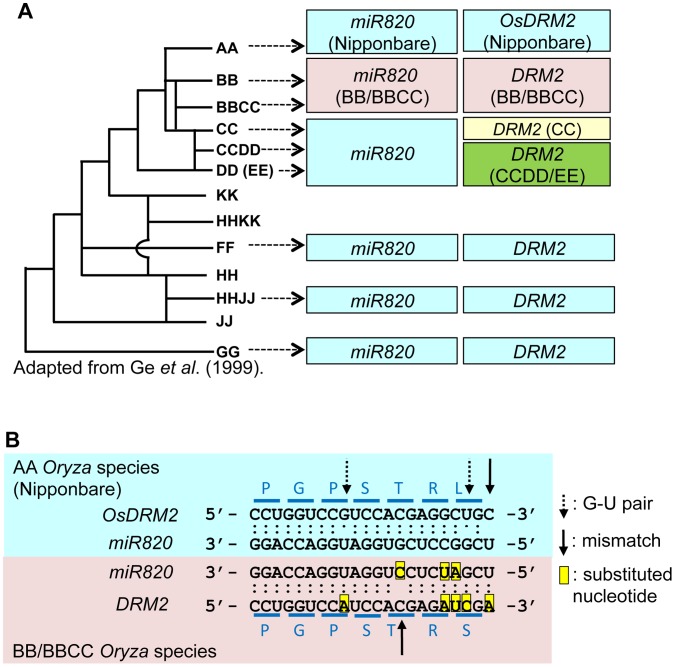
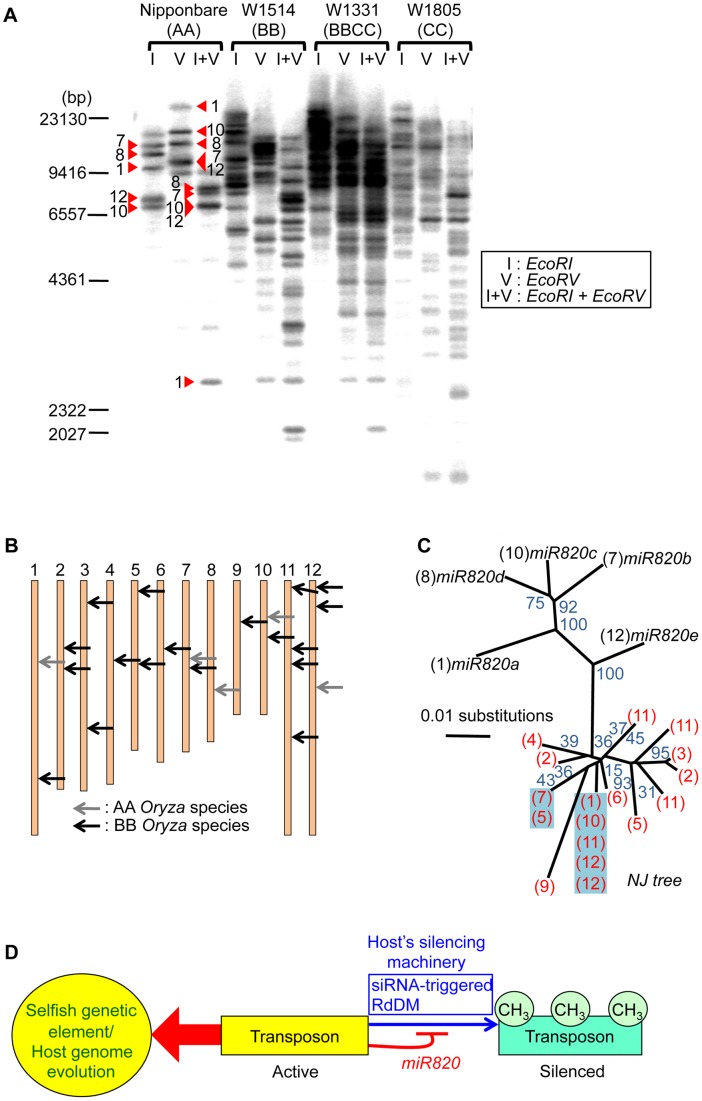
RNA silencing is a defense system against "genomic parasites" such as transposable elements (TE), which are potentially harmful to host genomes. In plants, transcripts from TEs induce production of double-stranded RNAs (dsRNAs) and are processed into small RNAs (small interfering RNAs, siRNAs) that suppress TEs by RNA-directed DNA methylation. Thus, the majority of TEs are epigenetically silenced. On the other hand, most of the eukaryotic genome is composed of TEs and their remnants, suggesting that TEs have evolved countermeasures against host-mediated silencing. Under some circumstances, TEs can become active and increase in copy number. Knowledge is accumulating on the mechanisms of TE silencing by the host; however, the mechanisms by which TEs counteract silencing are poorly understood. Here, we show that a class of TEs in rice produces a microRNA (miRNA) to suppress host silencing. Members of the microRNA820 (miR820) gene family are located within CACTA DNA transposons in rice and target a de novo DNA methyltransferase gene, OsDRM2, one of the components of epigenetic silencing. We confirmed that miR820 negatively regulates the expression of OsDRM2. In addition, we found that expression levels of various TEs are increased quite sensitively in response to decreased OsDRM2 expression and DNA methylation at TE loci. Furthermore, we found that the nucleotide sequence of miR820 and its recognition site within the target gene in some Oryza species have co-evolved to maintain their base-pairing ability. The co-evolution of these sequences provides evidence for the functionality of this regulation. Our results demonstrate how parasitic elements in the genome escape the host's defense machinery. Furthermore, our analysis of the regulation of OsDRM2 by miR820 sheds light on the action of transposon-derived small RNAs, not only as a defense mechanism for host genomes but also as a regulator of interactions between hosts and their parasitic elements.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The copy number of rice CACTA DNA transposons carrying MIR820 does not correlate with MIR820 expression.Plant Signal Behav. 2013 Aug;8(8):e25169. doi: 10.4161/psb.25169. Epub 2013 May 31. Plant Signal Behav. 2013. PMID: 23733074 Free PMC article.
-
Expression of the rice microRNA miR820 is associated with epigenetic modifications at its own locus.Genes Genet Syst. 2013;88(2):105-12. doi: 10.1266/ggs.88.105. Genes Genet Syst. 2013. PMID: 23832302
-
The initiation of epigenetic silencing of active transposable elements is triggered by RDR6 and 21-22 nucleotide small interfering RNAs.Plant Physiol. 2013 May;162(1):116-31. doi: 10.1104/pp.113.216481. Epub 2013 Mar 29. Plant Physiol. 2013. PMID: 23542151 Free PMC article.
-
How transposable elements are recognized and epigenetically silenced in plants?Curr Opin Plant Biol. 2023 Oct;75:102428. doi: 10.1016/j.pbi.2023.102428. Epub 2023 Jul 21. Curr Opin Plant Biol. 2023. PMID: 37481986 Review.
-
Transposable element origins of epigenetic gene regulation.Curr Opin Plant Biol. 2011 Apr;14(2):156-61. doi: 10.1016/j.pbi.2011.01.003. Epub 2011 Apr 4. Curr Opin Plant Biol. 2011. PMID: 21444239 Review.
Cited by
-
The copy number of rice CACTA DNA transposons carrying MIR820 does not correlate with MIR820 expression.Plant Signal Behav. 2013 Aug;8(8):e25169. doi: 10.4161/psb.25169. Epub 2013 May 31. Plant Signal Behav. 2013. PMID: 23733074 Free PMC article.
-
Control of transposable elements in Arabidopsis thaliana.Chromosome Res. 2014 Jun;22(2):217-23. doi: 10.1007/s10577-014-9417-9. Chromosome Res. 2014. PMID: 24801341 Review.
-
Host-transposon interactions: conflict, cooperation, and cooption.Genes Dev. 2019 Sep 1;33(17-18):1098-1116. doi: 10.1101/gad.327312.119. Genes Dev. 2019. PMID: 31481535 Free PMC article. Review.
-
The Evolutionary Volte-Face of Transposable Elements: From Harmful Jumping Genes to Major Drivers of Genetic Innovation.Cells. 2021 Oct 29;10(11):2952. doi: 10.3390/cells10112952. Cells. 2021. PMID: 34831175 Free PMC article. Review.
-
Dicer-like 3 produces transposable element-associated 24-nt siRNAs that control agricultural traits in rice.Proc Natl Acad Sci U S A. 2014 Mar 11;111(10):3877-82. doi: 10.1073/pnas.1318131111. Epub 2014 Feb 19. Proc Natl Acad Sci U S A. 2014. PMID: 24554078 Free PMC article.
References
-
- Plasterk RHA (2002) RNA silencing: The genome's immune system. Science 296: 1263–1265. - PubMed
-
- Almedia R, Allshire RC (2005) RNA silencing and genome regulation. Trends Cell Biol 15: 251–258. - PubMed
-
- Aravin AA, Hannon GJ, Brennecke J (2007) The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science 318: 761–764. - PubMed
-
- Saito K, Siomi MC (2010) Small RNA-mediated quiescence of transposable elements in animals. Dev Cell 19: 687–697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
