

The Relationship between fenestrations, sieve plates and rafts in liver sinusoidal endothelial cells
- PMID: 23029409
- PMCID: PMC3454341
- DOI: 10.1371/journal.pone.0046134
The Relationship between fenestrations, sieve plates and rafts in liver sinusoidal endothelial cells
Abstract
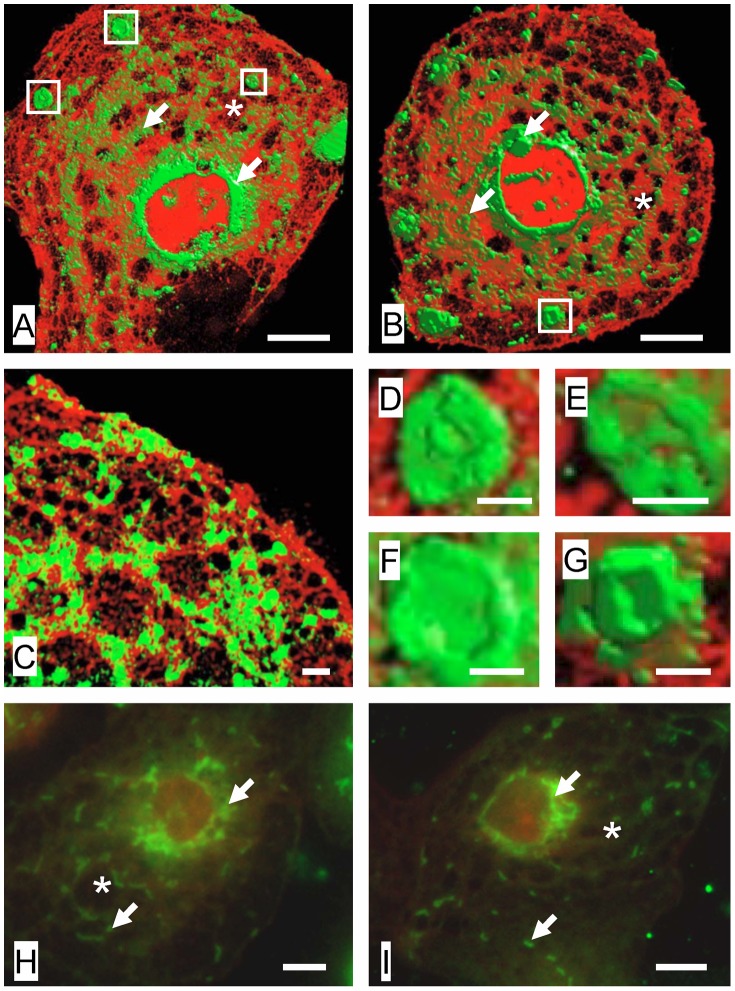
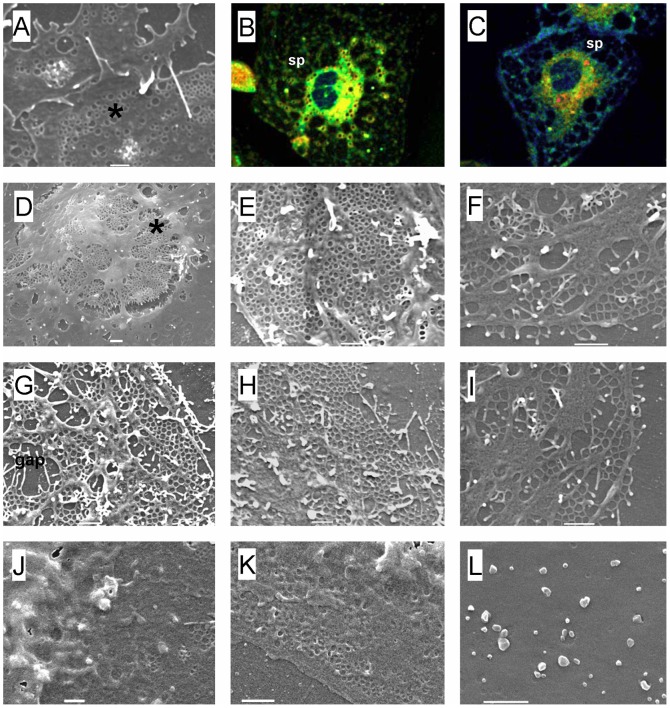
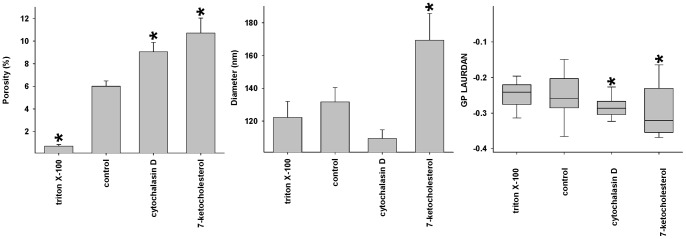
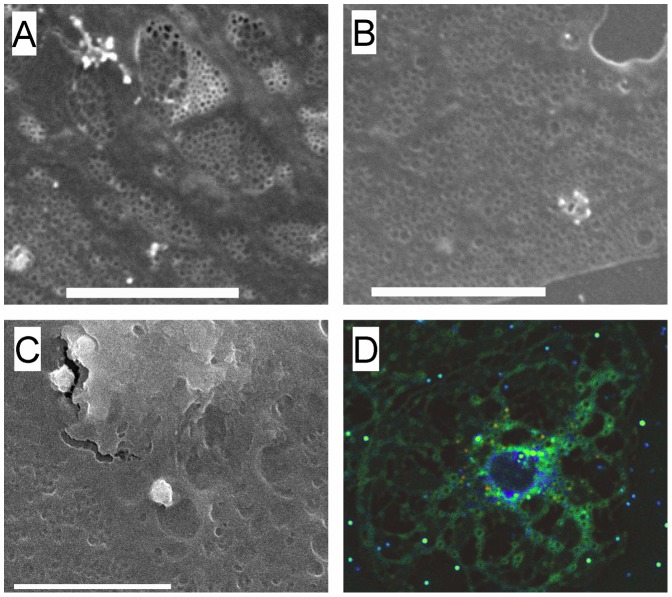
Fenestrations are transcellular pores in endothelial cells that facilitate transfer of substrates between blood and the extravascular compartment. In order to understand the regulation and formation of fenestrations, the relationship between membrane rafts and fenestrations was investigated in liver sinusoidal endothelial cells where fenestrations are grouped into sieve plates. Three dimensional structured illumination microscopy, scanning electron microscopy, internal reflectance fluorescence microscopy and two-photon fluorescence microscopy were used to study liver sinusoidal endothelial cells isolated from mice. There was an inverse distribution between sieve plates and membrane rafts visualized by structured illumination microscopy and the fluorescent raft stain, Bodipy FL C5 ganglioside GM1. 7-ketocholesterol and/or cytochalasin D increased both fenestrations and lipid-disordered membrane, while Triton X-100 decreased both fenestrations and lipid-disordered membrane. The effects of cytochalasin D on fenestrations were abrogated by co-administration of Triton X-100, suggesting that actin disruption increases fenestrations by its effects on membrane rafts. Vascular endothelial growth factor (VEGF) depleted lipid-ordered membrane and increased fenestrations. The results are consistent with a sieve-raft interaction, where fenestrations form in non-raft lipid-disordered regions of endothelial cells once the membrane-stabilizing effects of actin cytoskeleton and membrane rafts are diminished.
Conflict of interest statement
Figures
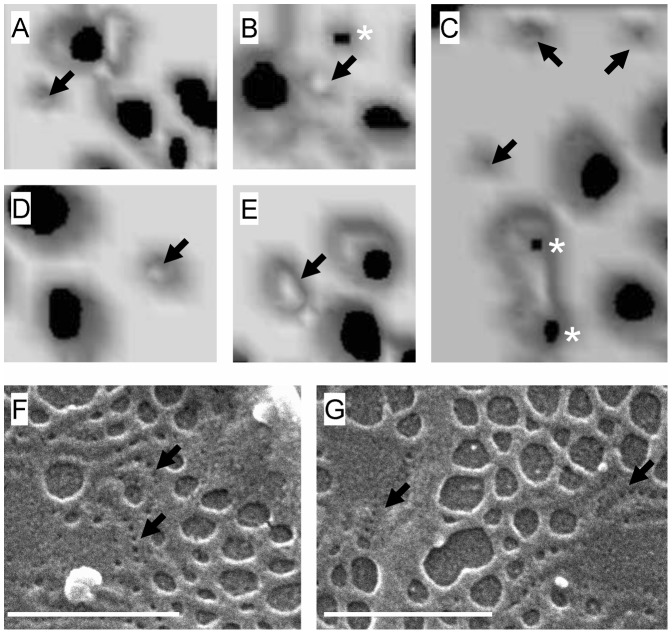
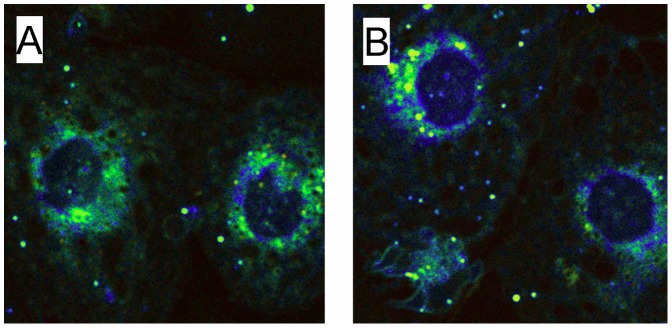
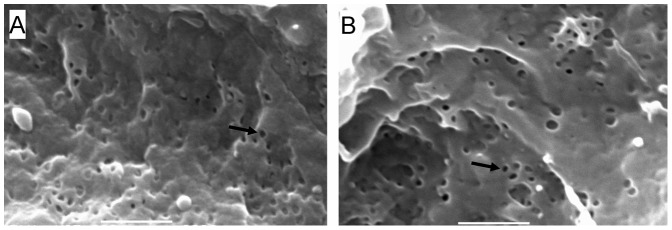
Comment in
-
Down the liver sinusoidal endothelial cell (LSEC) hole. Is there a role for lipid rafts in LSEC fenestration?Hepatology. 2013 Mar;57(3):1272-4. doi: 10.1002/hep.26249. Epub 2013 Feb 21. Hepatology. 2013. PMID: 23426851 No abstract available.
References
-
- Cogger VC, Le Couteur DG (2009) Fenestrations in the liver sinusoidal endothelial cell. In: Arias I, Wolkoff A, Boyer J, Shafritz D, Fausto N, et al., editors. The Liver: Biology and Pathobiology. 5th ed. Hokoben NJ: John Wiley & Sons, Ltd. 387–404.
-
- Hilmer SN, Cogger VC, Fraser R, McLean AJ, Sullivan D, et al. (2005) Age-related changes in the hepatic sinusoidal endothelium impede lipoprotein transfer in the rat. Hepatology 42: 1349–1354. - PubMed
-
- Le Couteur DG, Fraser R, Hilmer S, Rivory LP, McLean AJ (2005) The hepatic sinusoid in aging and cirrhosis - Effects on hepatic substrate disposition and drug clearance. Clin Pharmacokinet 44: 187–200. - PubMed
-
- Warren A, Le Couteur DG, Fraser R, Bowen DG, McCaughan GW, et al. (2006) T lymphocytes interact with hepatocytes through fenestrations in murine liver sinusoidal endothelial cells. Hepatology 44: 1182–1190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
