

Striatal adenosine signaling regulates EAAT2 and astrocytic AQP4 expression and alcohol drinking in mice
- PMID: 23032072
- PMCID: PMC3547194
- DOI: 10.1038/npp.2012.198
Striatal adenosine signaling regulates EAAT2 and astrocytic AQP4 expression and alcohol drinking in mice
Abstract
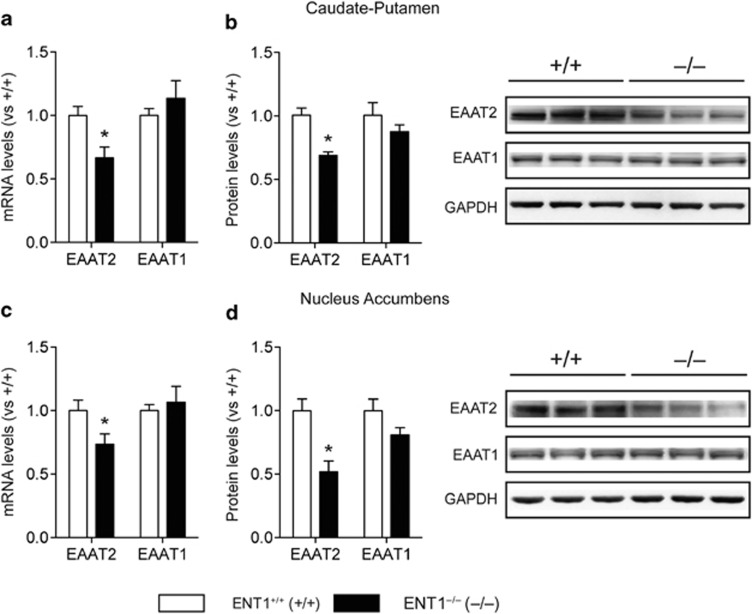
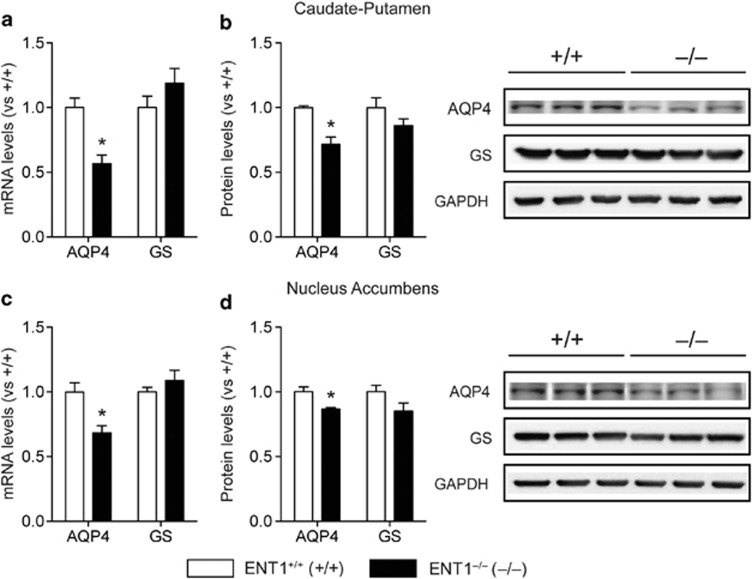
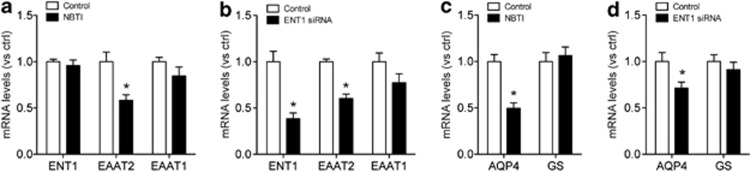
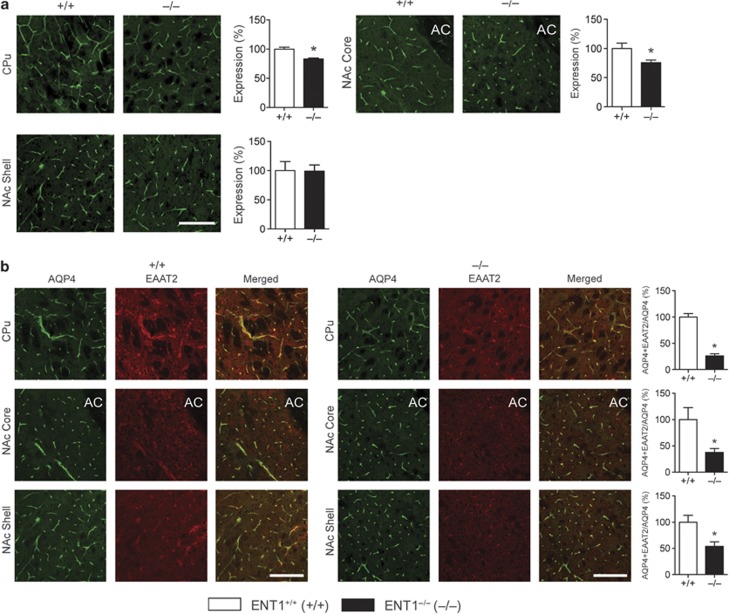
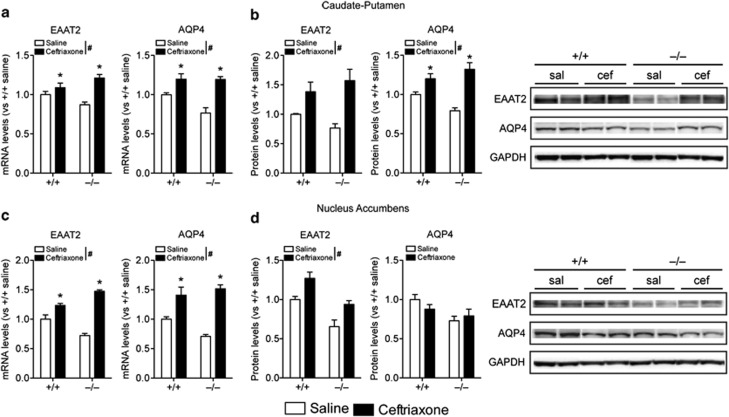
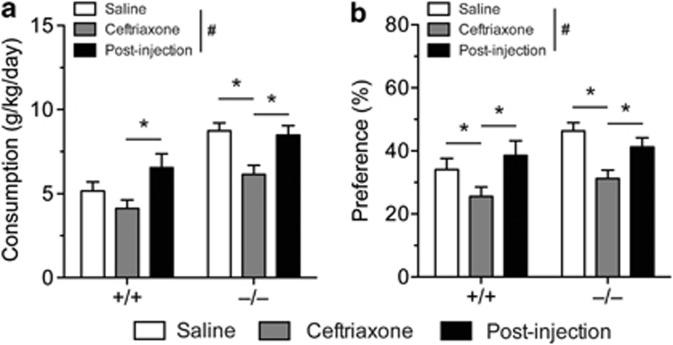
Adenosine signaling is implicated in several neuropsychiatric disorders, including alcoholism. Among its diverse functions in the brain, adenosine regulates glutamate release and has an essential role in ethanol sensitivity and preference. However, the molecular mechanisms underlying adenosine-mediated glutamate signaling in neuroglial interaction remain elusive. We have previously shown that mice lacking the ethanol-sensitive adenosine transporter, type 1 equilibrative nucleoside transporter (ENT1), drink more ethanol compared with wild-type mice and have elevated striatal glutamate levels. In addition, ENT1 inhibition or knockdown reduces glutamate transporter expression in cultured astrocytes. Here, we examined how adenosine signaling in astrocytes contributes to ethanol drinking. Inhibition or deletion of ENT1 reduced the expression of type 2 excitatory amino-acid transporter (EAAT2) and the astrocyte-specific water channel, aquaporin 4 (AQP4). EAAT2 and AQP4 colocalization was also reduced in the striatum of ENT1 null mice. Ceftriaxone, an antibiotic compound known to increase EAAT2 expression and function, elevated not only EAAT2 but also AQP4 expression in the striatum. Furthermore, ceftriaxone reduced ethanol drinking, suggesting that ENT1-mediated downregulation of EAAT2 and AQP4 expression contributes to excessive ethanol consumption in our mouse model. Overall, our findings indicate that adenosine signaling regulates EAAT2 and astrocytic AQP4 expressions, which control ethanol drinking in mice.
Figures
References
-
- Alliot F, Pessac B. Astrocytic cell clones derived from established cultures of 8-day postnatal mouse cerebella. Brain Res. 1984;306:283–291. - PubMed
-
- Brodie C, Kuperstein I, Acs P, Blumberg PM. Differential role of specific PKC isoforms in the proliferation of glial cells and the expression of the astrocytic markers GFAP and glutamine synthetase. Brain Res Mol Brain Res. 1998;56:108–117. - PubMed
-
- Choi DS, Cascini MG, Mailliard W, Young H, Paredes P, McMahon T, et al. The type 1 equilibrative nucleoside transporter regulates ethanol intoxication and preference. Nat Neurosci. 2004;7:855–861. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
