

Structural basis of efficient electron transport between photosynthetic membrane proteins and plastocyanin in spinach revealed using nuclear magnetic resonance
- PMID: 23032988
- PMCID: PMC3517244
- DOI: 10.1105/tpc.112.102517
Structural basis of efficient electron transport between photosynthetic membrane proteins and plastocyanin in spinach revealed using nuclear magnetic resonance
Abstract
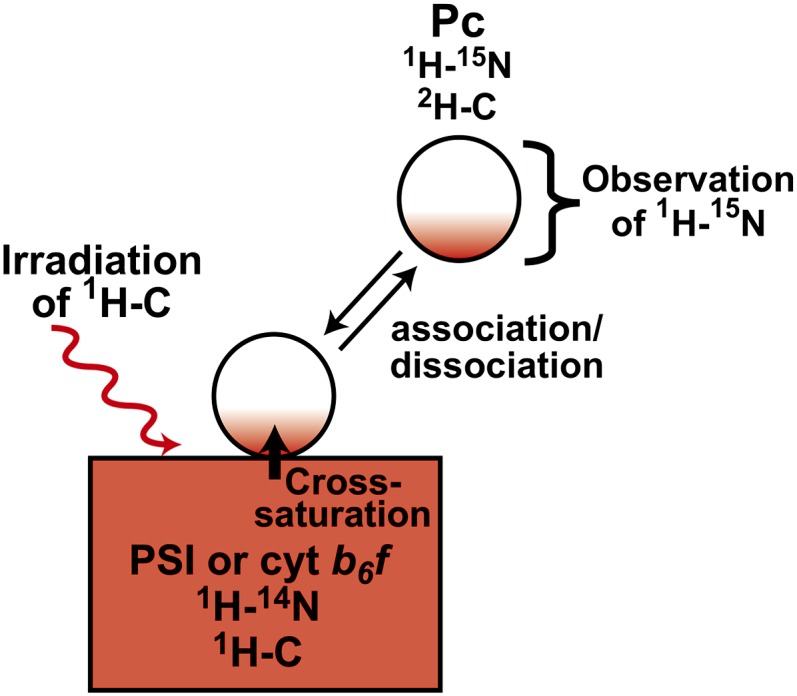
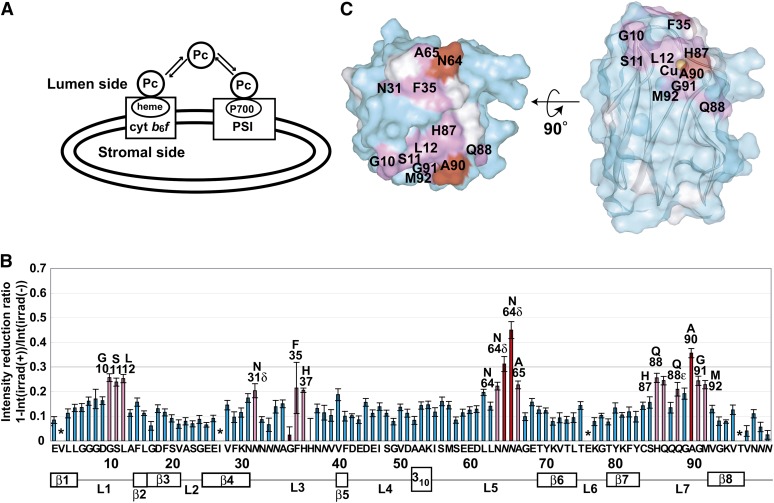
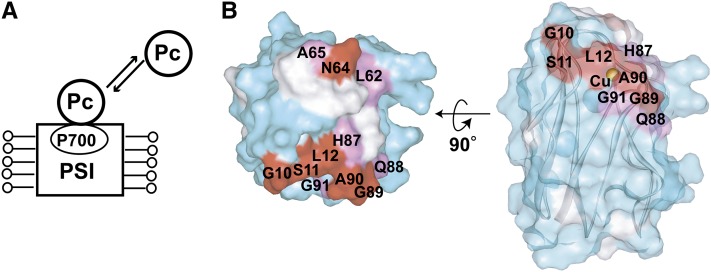
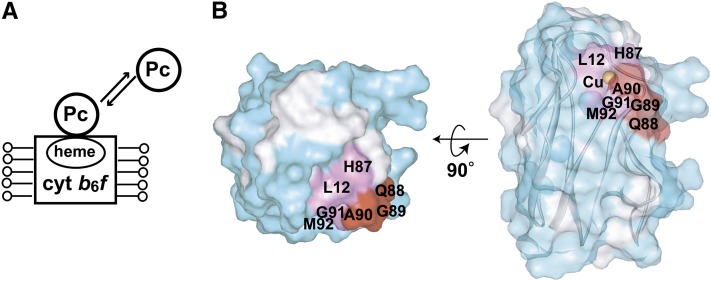
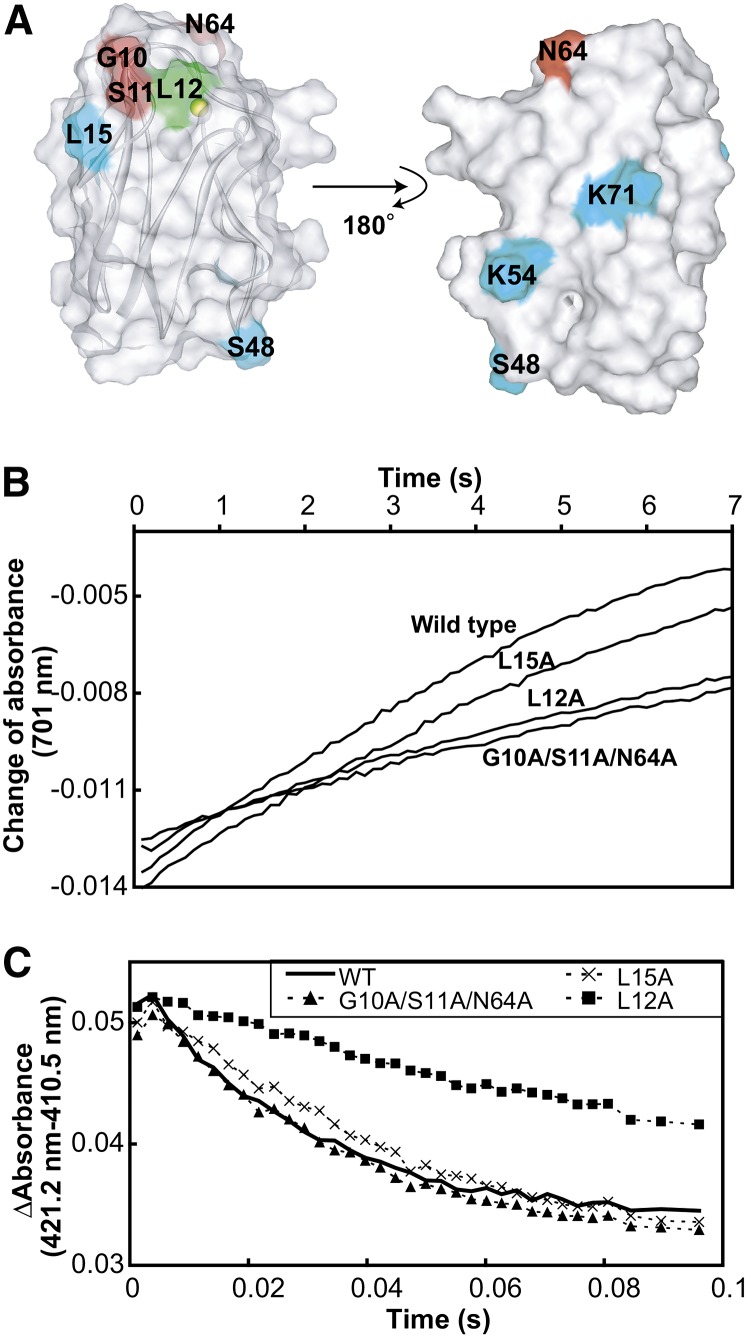
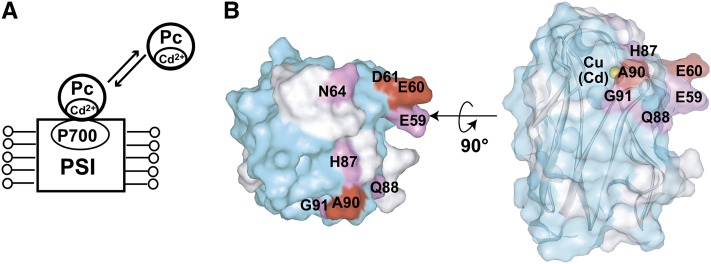
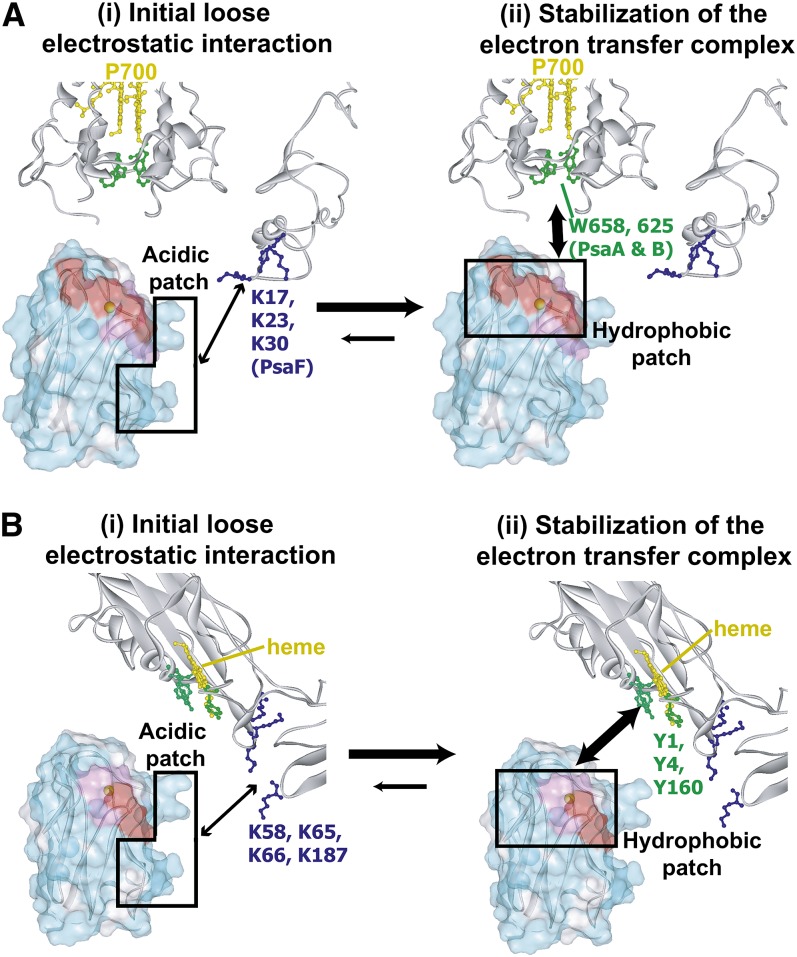
In the photosynthetic light reactions of plants and cyanobacteria, plastocyanin (Pc) plays a crucial role as an electron carrier and shuttle protein between two membrane protein complexes: cytochrome b(6)f (cyt b(6)f) and photosystem I (PSI). The rapid turnover of Pc between cyt b(6)f and PSI enables the efficient use of light energy. In the Pc-cyt b(6)f and Pc-PSI electron transfer complexes, the electron transfer reactions are accomplished within <10(-4) s. However, the mechanisms enabling the rapid association and dissociation of Pc are still unclear because of the lack of an appropriate method to study huge complexes with short lifetimes. Here, using the transferred cross-saturation method, we investigated the residues of spinach (Spinacia oleracea) Pc in close proximity to spinach PSI and cyt b(6)f, in both the thylakoid vesicle-embedded and solubilized states. We demonstrated that the hydrophobic patch residues of Pc are in close proximity to PSI and cyt b(6)f, whereas the acidic patch residues of Pc do not form stable salt bridges with either PSI or cyt b(6)f, in the electron transfer complexes. The transient characteristics of the interactions on the acidic patch facilitate the rapid association and dissociation of Pc.
Figures
References
-
- Amunts A., Ben-Shem A., Nelson N. (2005). Solving the structure of plant photosystem I—Biochemistry is vital. Photochem. Photobiol. Sci. 4: 1011–1015 - PubMed
-
- Amunts A., Drory O., Nelson N. (2007). The structure of a plant photosystem I supercomplex at 3.4 A resolution. Nature 447: 58–63 - PubMed
-
- Andersson B. (1986). Characterization of the thylakoid membrane by subfractionation analyses. Methods Enzymol. 118: 325–338
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
