

αB-Crystallin regulates expansion of CD11b⁺Gr-1⁺ immature myeloid cells during tumor progression
- PMID: 23033322
- PMCID: PMC3528311
- DOI: 10.1096/fj.12-213017
αB-Crystallin regulates expansion of CD11b⁺Gr-1⁺ immature myeloid cells during tumor progression
Abstract
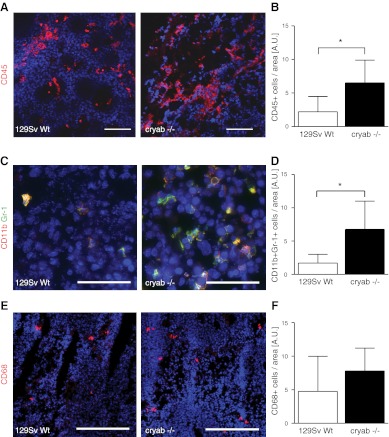
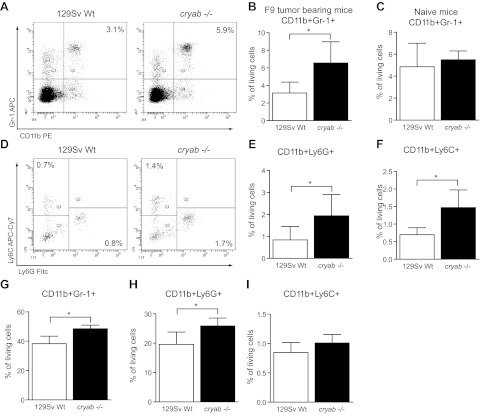
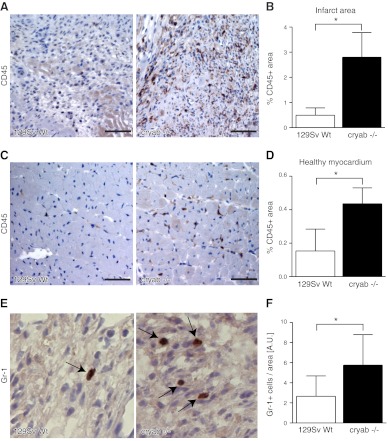
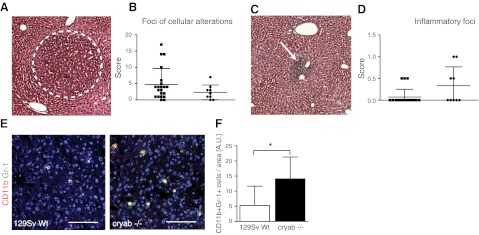
The molecular chaperone αB-crystallin has emerged as a target for cancer therapy due to its expression in human tumors and its role in regulating tumor angiogenesis. αB-crystallin also reduces neuroinflammation, but its role in other inflammatory conditions has not been investigated. Here, we examined whether αB-crystallin regulates inflammation associated with tumors and ischemia. We found that CD45(+) leukocyte infiltration is 3-fold increased in tumors and ischemic myocardium in αB-crystallin-deficient mice. Notably, αB-crystallin is prominently expressed in CD11b(+) Gr-1(+) immature myeloid cells (IMCs), known as regulators of angiogenesis and immune responses, while lymphocytes and mature granulocytes show low αB-crystallin expression. αB-Crystallin deficiency results in a 3-fold higher accumulation of CD11b(+) Gr-1(+) IMCs in tumors and a significant rise in CD11b(+) Gr-1(+) IMCs in spleen and bone marrow. Similarly, we noted a 2-fold increase in CD11b(+) Gr-1(+) IMCs in chronically inflamed livers in αB-crystallin-deficient mice. The effect of αB-crystallin on IMC accumulation is limited to pathological conditions, as CD11b(+) Gr-1(+) IMCs are not elevated in naive mice. Through ex vivo differentiation of CD11b(+) Gr-1(+) cells, we provide evidence that αB-crystallin regulates systemic expansion of IMCs through a cell-intrinsic mechanism. Our study suggests a key role of αB-crystallin in limiting expansion of CD11b(+) Gr-1(+) IMCs in diverse pathological conditions.
Figures

References
-
- Mantovani A., Allavena P., Sica A., Balkwill F. (2008) Cancer-related inflammation. Nature 454, 436–444 - PubMed
-
- Schafer M., Werner S. (2008) Cancer as an overhealing wound: an old hypothesis revisited. Nat. Rev. Mol. Cell. Biol. 9, 628–638 - PubMed
-
- Murdoch C., Muthana M., Coffelt S. B., Lewis C. E. (2008) The role of myeloid cells in the promotion of tumour angiogenesis. Nat. Rev. Cancer 8, 618–631 - PubMed
-
- Shojaei F., Wu X., Malik A. K., Zhong C., Baldwin M. E., Schanz S., Fuh G., Gerber H. P., Ferrara N. (2007) Tumor refractoriness to anti-VEGF treatment is mediated by CD11b+Gr1+ myeloid cells. Nat. Biotechnol. 25, 911–920 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
