

Inhibition of LpxC protects mice from resistant Acinetobacter baumannii by modulating inflammation and enhancing phagocytosis
- PMID: 23033474
- PMCID: PMC3518917
- DOI: 10.1128/mBio.00312-12
Inhibition of LpxC protects mice from resistant Acinetobacter baumannii by modulating inflammation and enhancing phagocytosis
Abstract
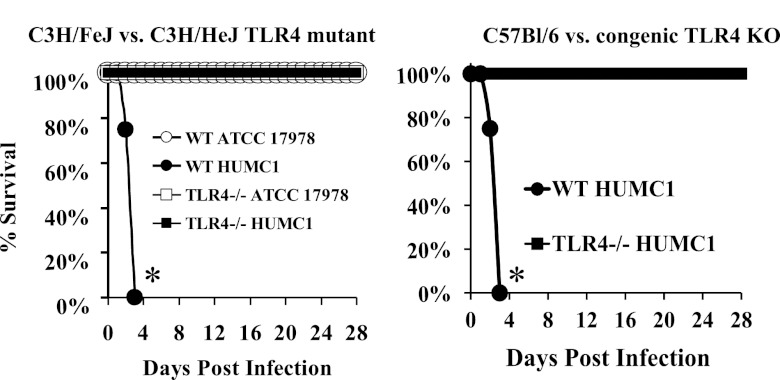
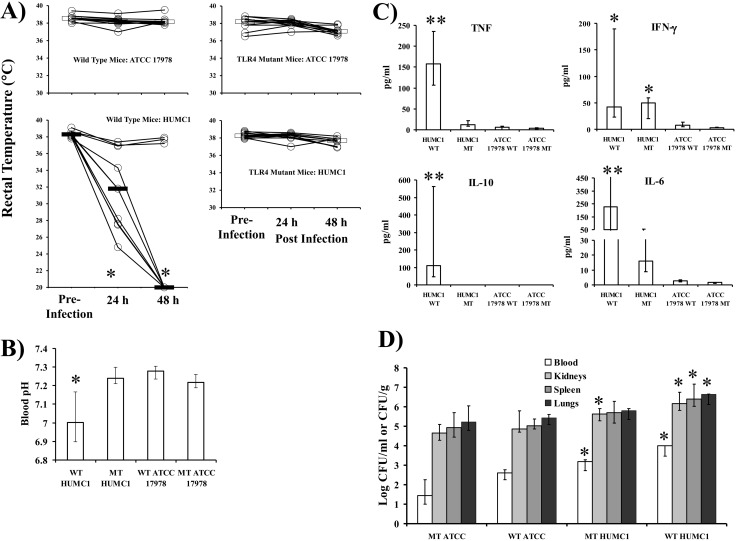
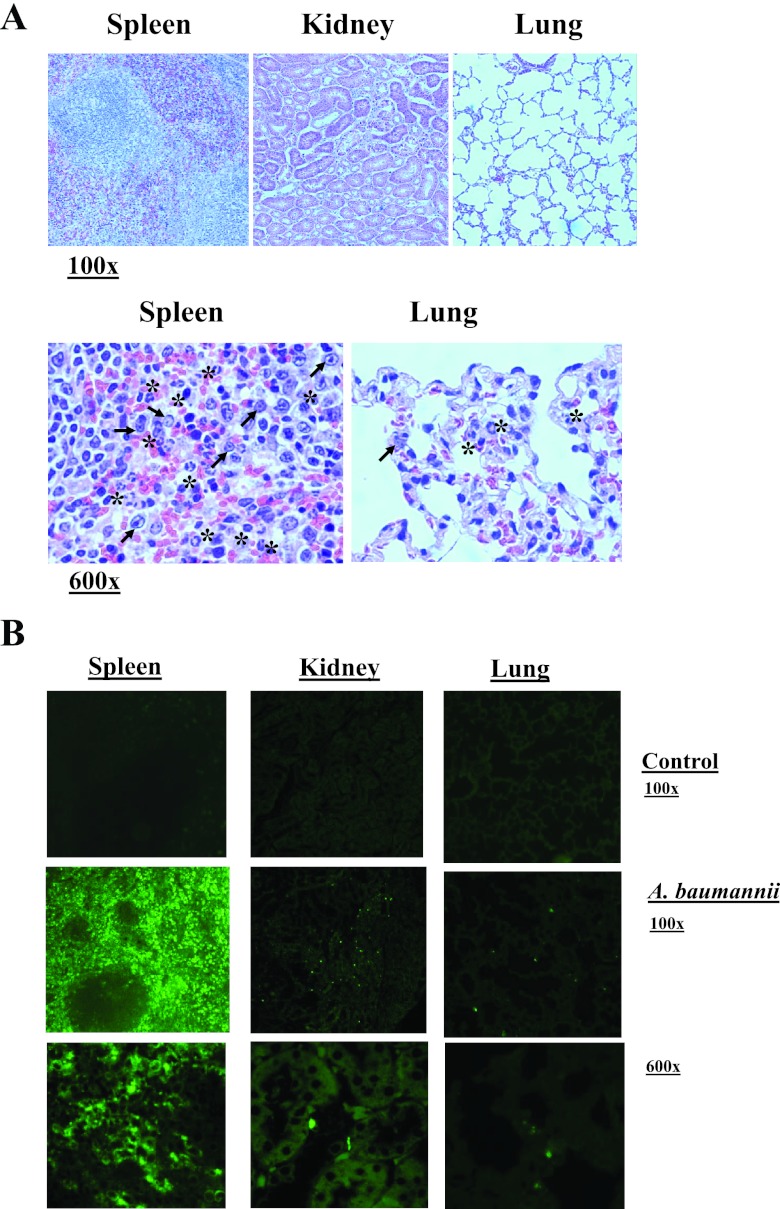
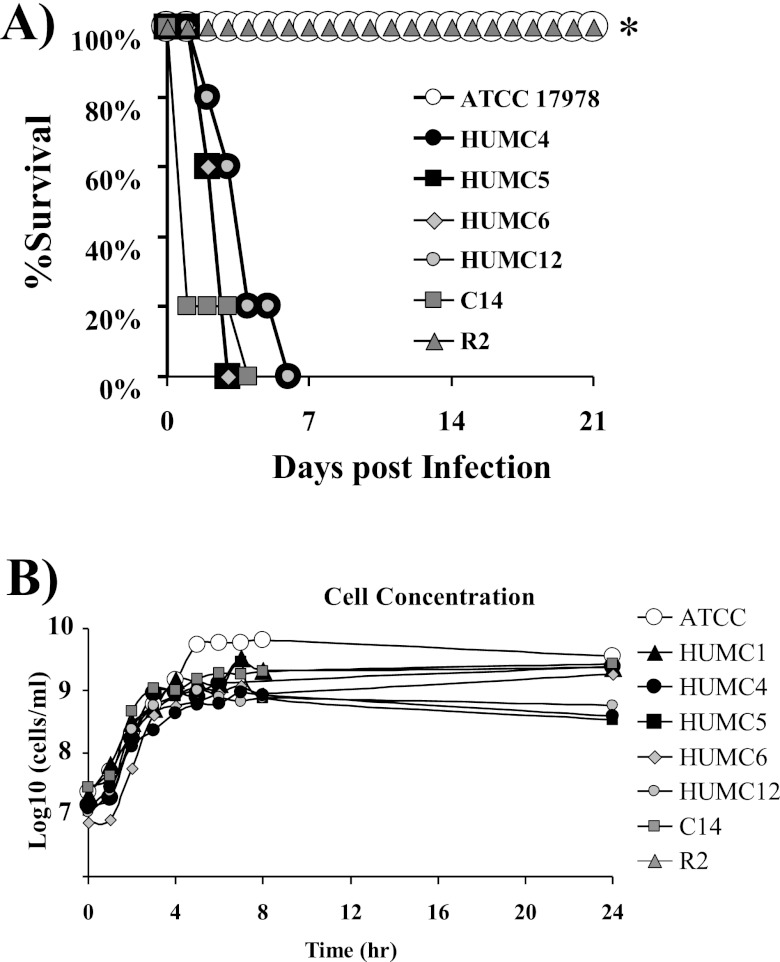
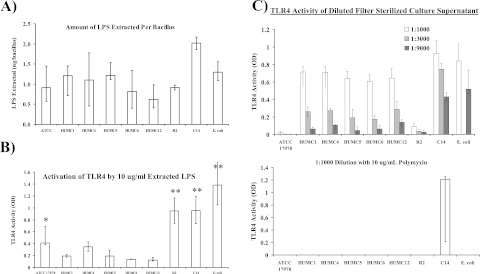
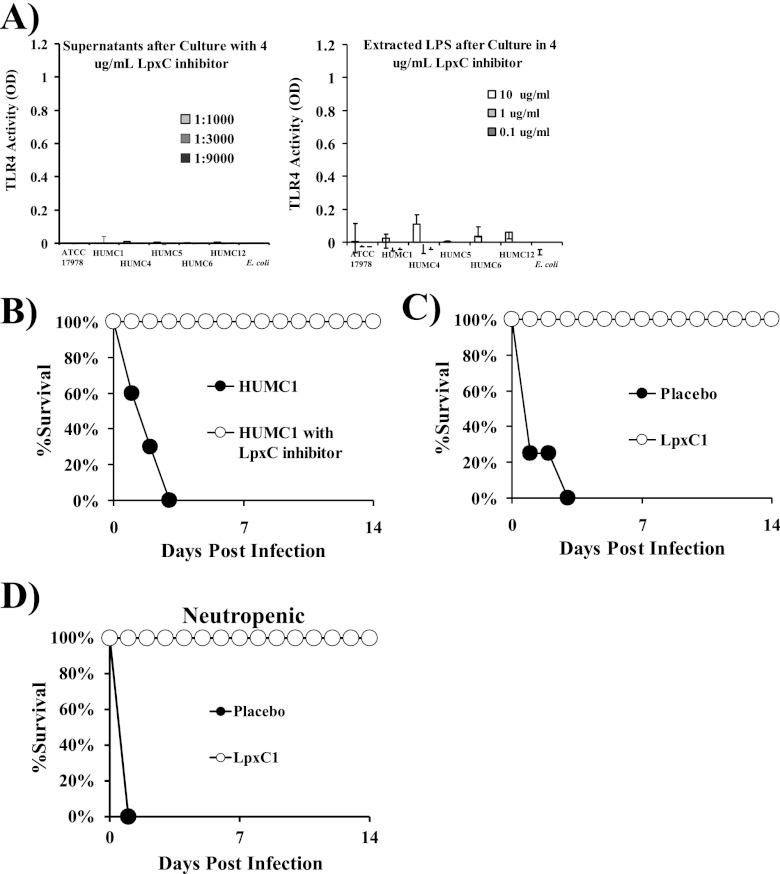
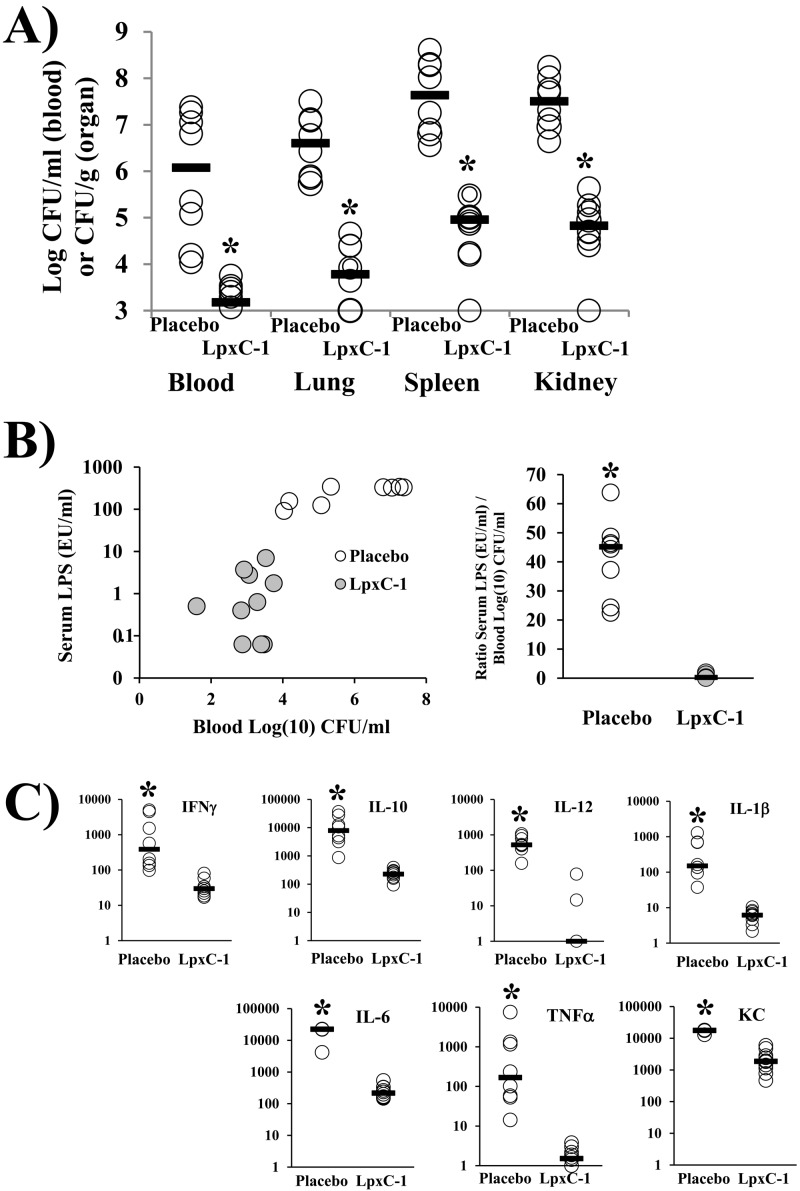
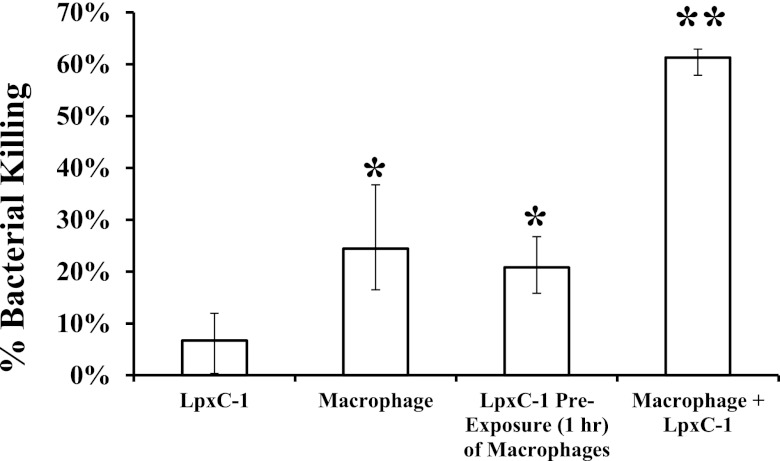
New treatments are needed for extensively drug-resistant (XDR) Gram-negative bacilli (GNB), such as Acinetobacter baumannii. Toll-like receptor 4 (TLR4) was previously reported to enhance bacterial clearance of GNB, including A. baumannii. However, here we have shown that 100% of wild-type mice versus 0% of TLR4-deficient mice died of septic shock due to A. baumannii infection, despite having similar tissue bacterial burdens. The strain lipopolysaccharide (LPS) content and TLR4 activation by extracted LPS did not correlate with in vivo virulence, nor did colistin resistance due to LPS phosphoethanolamine modification. However, more-virulent strains shed more LPS during growth than less-virulent strains, resulting in enhanced TLR4 activation. Due to the role of LPS in A. baumannii virulence, an LpxC inhibitor (which affects lipid A biosynthesis) antibiotic was tested. The LpxC inhibitor did not inhibit growth of the bacterium (MIC>512 µg/ml) but suppressed A. baumannii LPS-mediated activation of TLR4. Treatment of infected mice with the LpxC inhibitor enhanced clearance of the bacteria by enhancing opsonophagocytic killing, reduced serum LPS concentrations and inflammation, and completely protected the mice from lethal infection. These results identify a previously unappreciated potential for the new class of LpxC inhibitor antibiotics to treat XDR A. baumannii infections. Furthermore, they have far-reaching implications for pathogenesis and treatment of infections caused by GNB and for the discovery of novel antibiotics not detected by standard in vitro screens.
Importance: Novel treatments are needed for infections caused by Acinetobacter baumannii, a Gram-negative bacterium that is extremely antibiotic resistant. The current study was undertaken to understand the immunopathogenesis of these infections, as a basis for defining novel treatments. The primary strain characteristic that differentiated virulent from less-virulent strains was shedding of Gram-negative lipopolysaccharide (LPS) during growth. A novel class of antibiotics, called LpxC inhibitors, block LPS synthesis, but these drugs do not demonstrate the ability to kill A. baumannii in vitro. We found that an LpxC inhibitor blocked the ability of bacteria to activate the sepsis cascade, enhanced opsonophagocytic killing of the bacteria, and protected mice from lethal infection. Thus, an entire new class of antibiotics which is already in development has heretofore-unrecognized potential to treat A. baumannii infections. Furthermore, standard antibiotic screens based on in vitro killing failed to detect this treatment potential of LpxC inhibitors for A. baumannii infections.
Figures
References
-
- Chow JC, Young DW, Golenbock DT, Christ WJ, Gusovsky F. 1999. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 274:10689–10692 - PubMed
-
- Hoshino K, et al. 1999. Cutting edge: toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps gene product. J. Immunol. 162:3749–3752 - PubMed
-
- Poltorak A, et al. 1998. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282:2085–2088 - PubMed
-
- Bihl F, et al. 2003. Overexpression of Toll-like receptor 4 amplifies the host response to lipopolysaccharide and provides a survival advantage in transgenic mice. J. Immunol. 170:6141–6150 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
