

β-Catenin activates the HOXA10 and CDX4 genes in myeloid progenitor cells
- PMID: 23038246
- PMCID: PMC3501069
- DOI: 10.1074/jbc.M112.402172
β-Catenin activates the HOXA10 and CDX4 genes in myeloid progenitor cells
Abstract
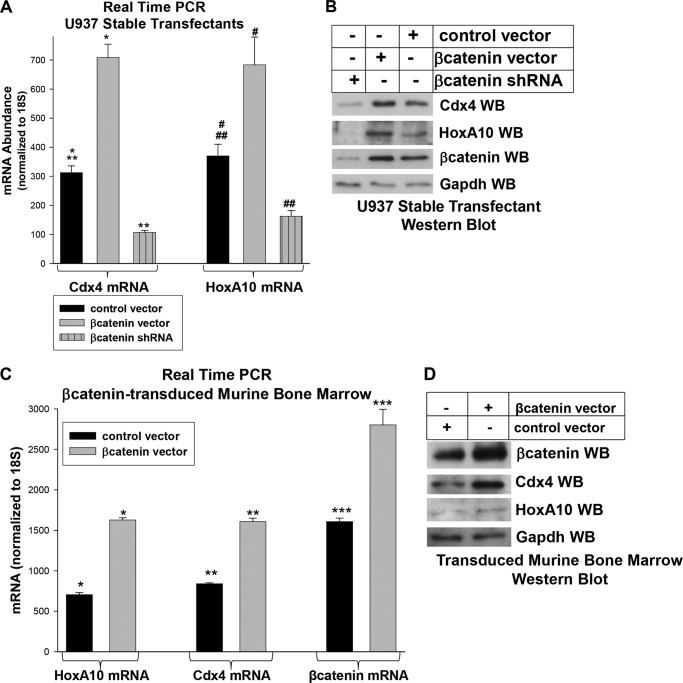
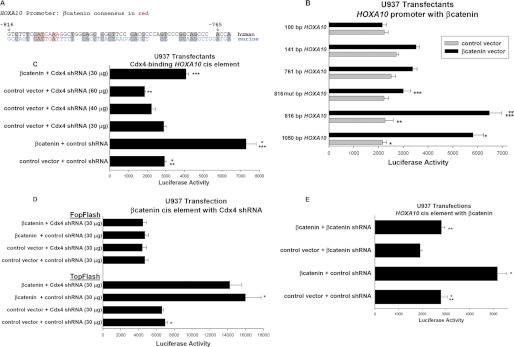
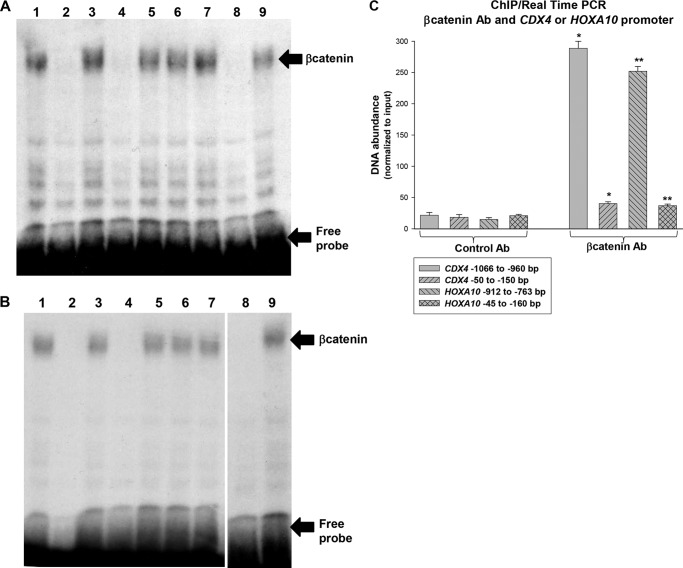
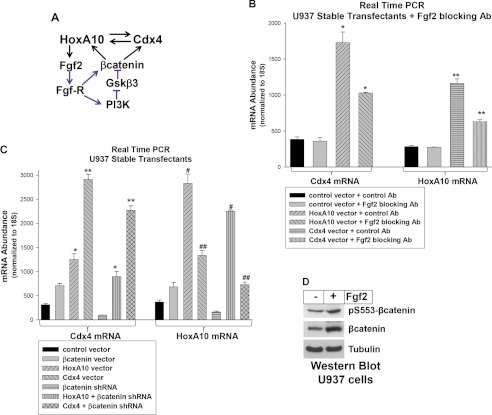
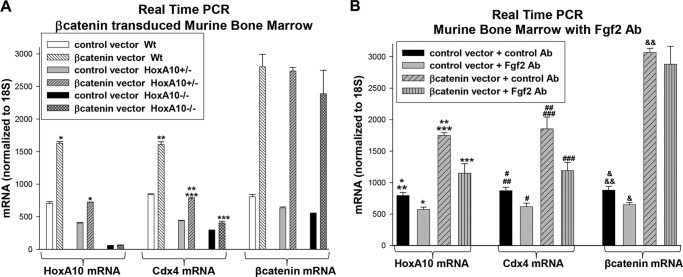
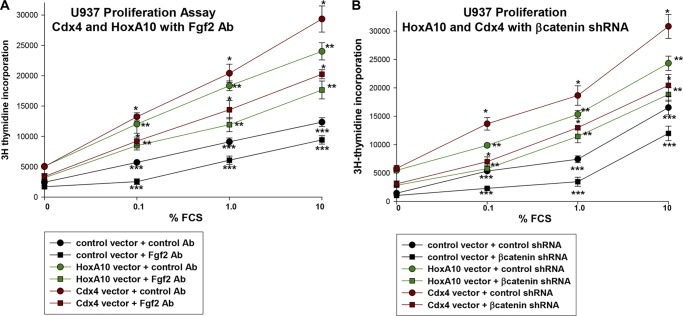
HoxA10 is a homeodomain transcription factor that is involved in maintenance of the myeloid progenitor population and implicated in myeloid leukemogenesis. Previously, we found that FGF2 and CDX4 are direct target genes of HoxA10 and that HOXA10 is a Cdx4 target gene. We also found that increased production of fibroblast growth factor 2 (Fgf2) by HoxA10-overexpressing myeloid progenitor cells results in activation of β-catenin in an autocrine manner. In this study, we identify novel cis elements in the CDX4 and HOXA10 genes that are activated by β-catenin in myeloid progenitor cells. We determine that β-catenin interacts with these cis elements, identifying both CDX4 and HOXA10 as β-catenin target genes in this context. We demonstrate that HoxA10-induced CDX4 transcription is influenced by Fgf2-dependent β-catenin activation. Similarly, Cdx4-induced HOXA10 transcription is influenced by β-catenin in an Fgf2-dependent manner. Increased expression of a set of Hox proteins, including HoxA10, is associated with poor prognosis in acute myeloid leukemia. Cdx4 contributes to leukemogenesis in Hox-overexpressing acute myeloid leukemia, and increased β-catenin activity is also associated with poor prognosis. The current studies identify a molecular mechanisms through which increased expression of HoxA10 increases Cdx4 expression by direct CDX4 activation and by Fgf2-induced β-catenin activity. This results in Cdx4-induced HoxA10-expression, creating a positive feedback mechanism.
Figures

Similar articles
-
Cooperation between AlphavBeta3 integrin and the fibroblast growth factor receptor enhances proliferation of Hox-overexpressing acute myeloid leukemia cells.Oncotarget. 2016 Aug 23;7(34):54782-54794. doi: 10.18632/oncotarget.10189. Oncotarget. 2016. PMID: 27340869 Free PMC article.
-
HoxA10 activates CDX4 transcription and Cdx4 activates HOXA10 transcription in myeloid cells.J Biol Chem. 2011 May 27;286(21):19047-64. doi: 10.1074/jbc.M110.213983. Epub 2011 Apr 6. J Biol Chem. 2011. PMID: 21471217 Free PMC article.
-
HoxA10 protein regulates transcription of gene encoding fibroblast growth factor 2 (FGF2) in myeloid cells.J Biol Chem. 2012 May 25;287(22):18230-48. doi: 10.1074/jbc.M111.328401. Epub 2012 Apr 4. J Biol Chem. 2012. PMID: 22493287 Free PMC article.
-
HOX gene regulation in acute myeloid leukemia: CDX marks the spot?Cell Cycle. 2007 Sep 15;6(18):2241-5. doi: 10.4161/cc.6.18.4656. Epub 2007 Jun 29. Cell Cycle. 2007. PMID: 17881901 Review.
-
The role of HOX genes in myeloid leukemogenesis.Curr Opin Hematol. 2006 Mar;13(2):67-73. doi: 10.1097/01.moh.0000208467.63861.d6. Curr Opin Hematol. 2006. PMID: 16456371 Review.
Cited by
-
Cooperation between AlphavBeta3 integrin and the fibroblast growth factor receptor enhances proliferation of Hox-overexpressing acute myeloid leukemia cells.Oncotarget. 2016 Aug 23;7(34):54782-54794. doi: 10.18632/oncotarget.10189. Oncotarget. 2016. PMID: 27340869 Free PMC article.
-
Downregulation of homeobox gene Barx2 increases gastric cancer proliferation and metastasis and predicts poor patient outcomes.Oncotarget. 2016 Sep 13;7(37):60593-60608. doi: 10.18632/oncotarget.11260. Oncotarget. 2016. PMID: 27533254 Free PMC article.
-
An aberrantly sustained emergency granulopoiesis response accelerates postchemotherapy relapse in MLL1-rearranged acute myeloid leukemia in mice.J Biol Chem. 2020 Jul 10;295(28):9663-9675. doi: 10.1074/jbc.RA120.013206. Epub 2020 May 28. J Biol Chem. 2020. PMID: 32467231 Free PMC article.
-
Consecutive epigenetically-active agent combinations act in ID1-RUNX3-TET2 and HOXA pathways for Flt3ITD+ve AML.Oncotarget. 2017 Dec 25;9(5):5703-5715. doi: 10.18632/oncotarget.23655. eCollection 2018 Jan 19. Oncotarget. 2017. PMID: 29464028 Free PMC article.
-
HOXA10 controls proliferation, migration and invasion in oral squamous cell carcinoma.Int J Clin Exp Pathol. 2015 Apr 1;8(4):3613-23. eCollection 2015. Int J Clin Exp Pathol. 2015. PMID: 26097543 Free PMC article.
References
-
- Sauvageau G., Thorsteinsdottir U., Eaves C. J., Lawrence H. J., Largman C., Lansdorp P. M., Humphries R. K. (1995) Overexpression of HOXB4 in hematopoietic cells causes the selective expansion of more primitive populations in vitro and in vivo. Genes Dev. 9, 1753–1765 - PubMed
-
- Calvo K. R., Sykes D. B., Pasillas M., Kamps M. P. (2000) Hoxa9 immortalizes a granulocyte-macrophage colony-stimulating factor-dependent promyelocyte capable of biphenotypic differentiation to neutrophils or macrophages, independent of enforced meis expression. Mol. Cell. Biol. 20, 3274–3285 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
