

Human β cell transcriptome analysis uncovers lncRNAs that are tissue-specific, dynamically regulated, and abnormally expressed in type 2 diabetes
- PMID: 23040067
- PMCID: PMC3475176
- DOI: 10.1016/j.cmet.2012.08.010
Human β cell transcriptome analysis uncovers lncRNAs that are tissue-specific, dynamically regulated, and abnormally expressed in type 2 diabetes
Abstract
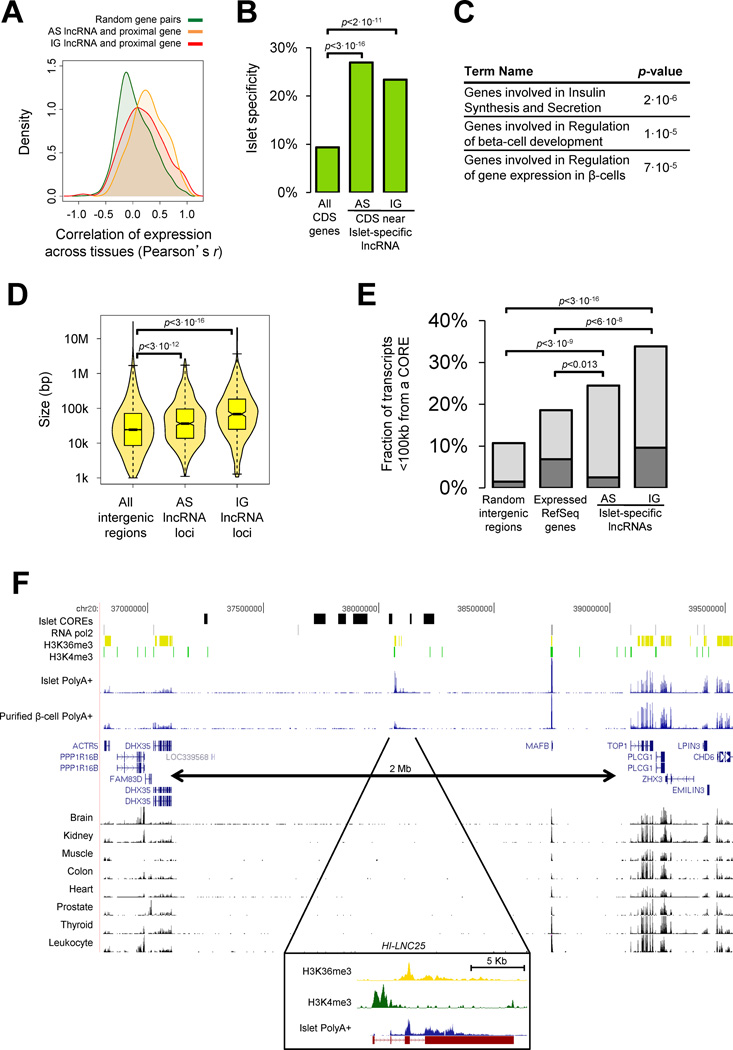
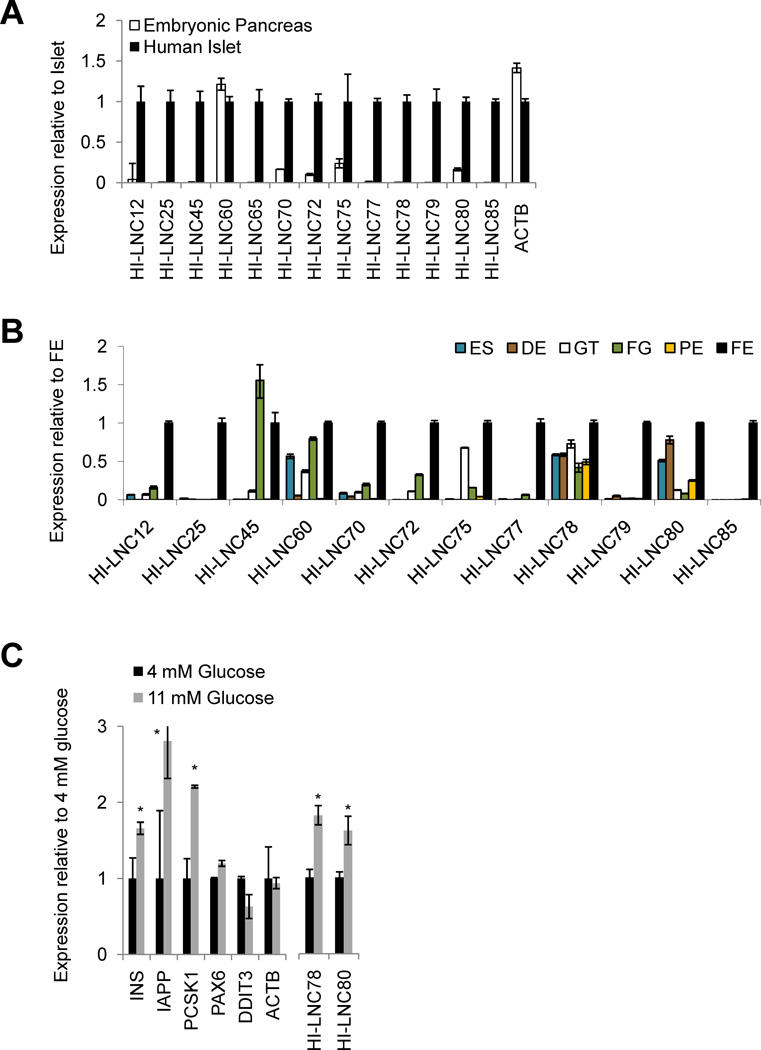
A significant portion of the genome is transcribed as long noncoding RNAs (lncRNAs), several of which are known to control gene expression. The repertoire and regulation of lncRNAs in disease-relevant tissues, however, has not been systematically explored. We report a comprehensive strand-specific transcriptome map of human pancreatic islets and β cells, and uncover >1100 intergenic and antisense islet-cell lncRNA genes. We find islet lncRNAs that are dynamically regulated and show that they are an integral component of the β cell differentiation and maturation program. We sequenced the mouse islet transcriptome and identify lncRNA orthologs that are regulated like their human counterparts. Depletion of HI-LNC25, a β cell-specific lncRNA, downregulated GLIS3 mRNA, thus exemplifying a gene regulatory function of islet lncRNAs. Finally, selected islet lncRNAs were dysregulated in type 2 diabetes or mapped to genetic loci underlying diabetes susceptibility. These findings reveal a new class of islet-cell genes relevant to β cell programming and diabetes pathophysiology.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures





References
-
- Bell GI, Polonsky KS. Diabetes mellitus and genetically programmed defects in beta-cell function. Nature. 2001;414:788–791. - PubMed
-
- Bensellam M, Van Lommel L, Overbergh L, Schuit FC, Jonas JC. Cluster analysis of rat pancreatic islet gene mRNA levels after culture in low-, intermediate- and high-glucose concentrations. Diabetologia. 2009;52:463–476. - PubMed
-
- Bertone P, Stolc V, Royce TE, Rozowsky JS, Urban AE, Zhu X, Rinn JL, Tongprasit W, Samanta M, Weissman S, et al. Global identification of human transcribed sequences with genome tiling arrays. Science. 2004;306:2242–2246. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
